來源簡介
人胚胎腎臟成纖維293細(xì)胞,通常指的是HEK 293,HEK-293,293 cells,都是從1970s的一個受孕女性胚胎中分離得到的永生化細(xì)胞系[1]。
1973年,來自荷蘭,萊頓的Alex van der Eb實(shí)驗(yàn)室,第一次將腺病毒DNA轉(zhuǎn)染進(jìn)體外培養(yǎng)的293細(xì)胞中。1977年,F(xiàn)rank Graham在McMaster大學(xué)發(fā)表了HEK轉(zhuǎn)染的文獻(xiàn),因?yàn)榍『肏EK細(xì)胞已經(jīng)傳代了293次,所以就命名為HEK293細(xì)胞。
一項(xiàng)關(guān)于HEK 293細(xì)胞系及其五個衍生細(xì)胞系的基因組和轉(zhuǎn)錄組的全面研究,將HEK 293的轉(zhuǎn)錄組與人類腎臟、腎上腺、垂體和中樞神經(jīng)組織的轉(zhuǎn)錄組進(jìn)行了比較。發(fā)現(xiàn)該細(xì)胞的性質(zhì)與腎上腺更接近[2]。

表達(dá)優(yōu)勢
經(jīng)過多年的發(fā)展,來源于腎臟的HEK293細(xì)胞發(fā)展成,廣泛應(yīng)用的成纖維型,內(nèi)皮型和上皮型細(xì)胞。細(xì)胞形態(tài)多為三角形或梭形,見圖1。
使用腺病毒感染HEK293細(xì)胞,經(jīng)過測序驗(yàn)證發(fā)現(xiàn)腺病毒約4.5kbp的左臂基因組,整合到了細(xì)胞的19號染色體上[3]。由此,使用腺病毒攜帶不同基因,對HEK293細(xì)胞進(jìn)行改造,生成了適應(yīng)不同藥物的多種細(xì)胞亞株。
經(jīng)過十年的發(fā)展,目前工業(yè)與科研界已經(jīng)進(jìn)化出了,293f,293T和293S等類型。見圖2。如上述提到的,在1973年,HEK293,可以感染Ad5腺病毒,篩選永生化細(xì)胞;在1984年,HEK293S能夠?qū)崿F(xiàn)MEN培養(yǎng)基的懸浮生長;在1985年,篩選到溫感的SV40 T細(xì)胞大抗原細(xì)胞系,增加了SV40起始子的表達(dá)量;1993年的HEK293H,該細(xì)胞貼壁狀態(tài)下,生長迅速,蛋白表達(dá)水平高,適應(yīng)了serum-free medium;隨后,又衍生出了HEK293FT,一種能夠感染慢病毒的neo抗性突變的細(xì)胞;2014年,優(yōu)化出適合商業(yè)化大規(guī)模培養(yǎng)的HEK293F;期間,HEK293E(EBVA)和HEK2936E,前者能夠表達(dá)EBNA-1蛋白,后者能夠表達(dá)Gly-Gly-Ala重復(fù)區(qū)域缺失的EBNA1t蛋白;還有2001年的HEK293FTM,含有穩(wěn)定轉(zhuǎn)染的FRT位點(diǎn)和TetR質(zhì)粒;還有2002年的HEK293SG,2010年的HEK293SGGD,HEK293A,HEK293MSR等適應(yīng)不同功能和藥物的改造細(xì)胞系。其修飾的靶基因與包括細(xì)胞增殖、凋亡、中心碳代謝、聚糖譜均一性、糖基化效率、蛋白質(zhì)折疊、分泌系統(tǒng)、蛋白質(zhì)表達(dá)等功能相關(guān)。
目前哺乳動物表達(dá)系統(tǒng)應(yīng)用最成熟,最廣泛的仍然是CHO表達(dá)系統(tǒng)。但是對于快速瞬轉(zhuǎn),人物種翻譯后修飾相似性,還有大分子蛋白復(fù)合物顆粒的表達(dá),293系統(tǒng)有著核心的優(yōu)勢。所以,293系統(tǒng)仍然是最有應(yīng)用潛力的哺乳動物表達(dá)系統(tǒng)之一。與CHO的表達(dá)優(yōu)勢比較,見下表1:
|
對比維度 |
HEK 293 細(xì)胞 |
CHO 細(xì)胞 |
|
核心定位 |
快速研發(fā)與復(fù)雜蛋白生產(chǎn) |
規(guī)模化生產(chǎn)的金標(biāo)準(zhǔn) |
|
細(xì)胞來源 |
人源 |
小鼠 |
|
核心優(yōu)勢 |
人物種翻譯后修飾(糖基化、γ-羧化等) 生產(chǎn)病毒載體,膜蛋白,多亞基復(fù)合物 |
表達(dá)產(chǎn)量極高,工藝成熟, 可穩(wěn)定放大,糖基化定向優(yōu)化 |
|
劣勢 |
1. 穩(wěn)定株產(chǎn)量低于CHO,大規(guī)模生產(chǎn)成本較高 2. 懸浮馴化/克隆篩選平臺不如CHO成熟 3. 攜帶腺病毒基因片段(E1),需安全性評估 |
1. 翻譯后修飾非完全人源化,可能產(chǎn)生免疫原性糖型(如NGNA) 2. 瞬時表達(dá)效率低,早期研發(fā)速度慢 3. 對某些復(fù)雜人源蛋白(尤其膜蛋白)表達(dá)困難或活性低 |
|
應(yīng)用范圍 |
膜蛋白/受體(GPCRs) AAV/慢病毒/凝血因子 |
商業(yè)化抗體藥/Fc融合蛋白/激素/細(xì)胞因子 |
|
流程 |
瞬時轉(zhuǎn)染為主→1-2周獲得蛋白樣品→若需生產(chǎn),再開發(fā)穩(wěn)定細(xì)胞株。 |
穩(wěn)定細(xì)胞株開發(fā)為主→耗時數(shù)月篩選高表達(dá)克隆→進(jìn)行工藝開發(fā)與放大。 |
表1
基因改造
因?yàn)?93的核心優(yōu)勢,科研與工業(yè)界不斷對293基因組進(jìn)行改造,主要關(guān)注細(xì)胞凋亡,細(xì)胞增殖,細(xì)胞中心碳代謝,糖基化,蛋白折疊,蛋白分泌等功能。借助的技術(shù)手段主要是通過miRNA和siRNA為主的敲除和敲降抑制,進(jìn)行蛋白組學(xué)分析,做合適表達(dá)條件的高通量篩選,見下圖3。

圖3
基因功能,如Cyclin-dependent kinase like 3 (CDKL3)基因,能夠增強(qiáng)蛋白表達(dá)的,促進(jìn)G1-S轉(zhuǎn)化的細(xì)胞周期基因和促進(jìn)合成功能的Cytochrome c oxidase subunit 15 (COX15)基因。還有一些細(xì)胞生長類的基因,如Cyclin-dependent Kinase inhibitor 2C (CDKN2C),也是調(diào)節(jié)G1進(jìn)程;抑制細(xì)胞凋亡蛋白合成的B-cell lymphoma 2 (BCL2)基因;降低脫氫酶激酶活性,并增加細(xì)胞質(zhì)中丙酮酸向乳酸的轉(zhuǎn)化的Dehydrogenase kinase (PDK)。除此之外,還有敲除高甘露糖修飾的MGAT1基因。其中高頻的甘露糖修飾,會增加藥物的那白被修飾的風(fēng)險,且更容易被呈遞給免疫細(xì)胞,增加藥物的體內(nèi)清除可能。相關(guān)基因類型,如下表2所示。
|
Functions |
Gene |
|
Proliferation |
CDKL3, COX15, EIF3I, CDKN1A/B, CDKN2C, FGF1 |
|
Apoptosis |
XIAP, BCL2, CASP3/6/7, AIF1, BAX/BAK, CrmA, NRF2 |
|
Central carbon metabolism |
Carboxylase, Dehydrogenase, PDK, HIF1A |
|
Glycan pattern homogeneity Glycosylation efficiency |
EndoT8/β-galactoside-α-2,6-sialyltransferase 1, MAN1A1/2, MAN1B1, FUT8,MAN1C1/MGAT1, Sialyl transferases, ST6GAL1, ST3GAL3/4, MAN2A1/2, |
|
Protein folding |
RIC3, XBP1, PDIA2, Hsp70/HSPA5, VKORC1, CALU |
|
Secretory machinery |
SNAP-23, VAMP8, MUNC18, sly1, Tetraspanin CD9 |
|
Protein expression |
HIPK1, miRNA-22-3b, CEBPG, OAZ1, CASP8AP2, FABP5, RETNLB, PC, XBP1, HSPA5, HERPUD1, NEDD8/4L, UGCG, CIT, CNP |
表2
基于上述HEK293的功能和應(yīng)用潛力,賽唐生物開發(fā)了HEK293 HCP ELISA(HH-H0019-3A1)試劑盒。該試劑盒的DL與QL可以控制在0.39ng/mL和0.78ng/mL。線性表現(xiàn)優(yōu)異80-120%,已經(jīng)證實(shí)可適用樣本為重組單抗,雙抗和病毒包裝樣本。數(shù)據(jù)見下表3:
|
QC |
Data source(2025-2026) |
|||||
|
DL |
0.39ng/mL |
|||||
|
QL |
0.78-800ng/mL |
|||||
|
Linear |
70%-130% |
Diluent |
Mean OD |
ng/mL |
Con ng/mL |
Recovery |
|
40× |
0.33 |
91.8 |
3670.4 |
96.32% |
||
|
80× |
0.168 |
41.0 |
3279.0 |
89.34% |
||
|
160× |
0.098 |
21.0 |
3364.2 |
102.60% |
||
|
320× |
0.063 |
11.8 |
3767.4 |
111.99% |
||
|
640× |
0.036 |
5.2 |
3312.7 |
87.93% |
||
|
Recovery |
70%-130%(8×Buffer) |
|||||
|
CV(S2/S4/S6) |
<3.0%-4.6% |
|||||
|
Samples |
mAb/BiAb/AAV |
|||||
表3
同時比較了Cygnus產(chǎn)品,貨號: F650S。在(重組蛋白類型)不同藥物(相同藥物不同純化階段)中的檢出率,有不錯的可比性,見下圖4,5。
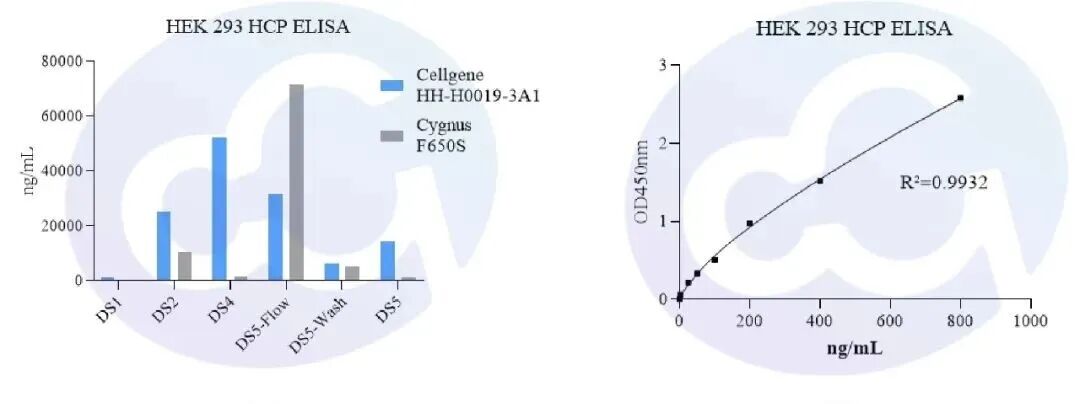
圖4 圖5
同時,在病毒包裝樣本中,從收獲液,澄清液和病毒濃縮液樣本中,多數(shù)高于Cygnus F650S,并遠(yuǎn)高于國內(nèi)平行產(chǎn)品的檢測結(jié)果。通過ELISA進(jìn)一步驗(yàn)證了客戶HCP清除工藝效率,符合客戶多年純化工藝積淀數(shù)據(jù)。且由于終產(chǎn)物中的HCP識別純度表現(xiàn)優(yōu)異,更容易通過藥監(jiān)局審批。見圖6-9。

圖6 圖7

圖8 圖9
另外,賽唐生物也做了充分的特異性檢測,如核酸酶,胰蛋白酶,BSA,HCP等,極大程度上排除了非特異性識別,見下表4:
|
Samples |
Con ng/mL |
OD |
ng/mL |
Cross-reaction |
|
Nuclease |
1000 |
0.013 |
0 |
0.00% |
|
800 |
0.0117 |
0 |
0.00% |
|
|
Trypsin |
250 |
0.0205 |
0 |
0.00% |
|
125 |
0.014 |
0 |
0.00% |
|
|
Bovine Serum Albumin |
32 |
0.0127 |
0 |
0.00% |
|
CHO HCP |
200 |
0.0213 |
0 |
0.00% |
|
E.coli HCP |
100 |
0.0143 |
0 |
0.00% |
|
SF9 |
400 |
0.017 |
0 |
0.00% |
|
Hansenula polymorpha HCP |
800 |
0.0825 |
0 |
0.00% |
|
Pichia pastori X-33 HCP |
800 |
0.0195 |
0 |
0.00% |
|
Saccharomyces cerevisiae HCP |
800 |
0.072 |
0 |
0.00% |
|
Pichia pastori GS115 HCP |
100 |
0.016 |
0 |
0.00% |
|
Human IgM |
50 |
0.011 |
0 |
0.00% |
|
Human IgG |
5 |
0.0115 |
0 |
0.00% |
表4
賽唐生物始終堅(jiān)持以解決實(shí)際問題導(dǎo)向,和藥企客戶踏實(shí)走好每一步。“權(quán)威”無法代替技術(shù)去解決實(shí)際問題,“金標(biāo)準(zhǔn)”不是唯一,但更要和藥企一起堅(jiān)持多年的法規(guī)積累,ELISA與LC-MS,二者缺一不可。解決不了ELISA問題,遑論解決以此為基礎(chǔ)的LC-MS鑒定問題?腳下懸空很危險,非此即彼不可取。
參考文獻(xiàn):
[1] Kavsan, Vadym M; et al. Immortalized cells and one oncogene in malignant transformation: old insights on new explanation. BMC Cell Biology. 2011. 12: 23.
[2] Yao-Cheng Lin, et al. Genome dynamics of the human embryonic kidney 293 lineage in response to cell biology manipulations. Nature communications. 2014. 5:4767.
[3] Louis N, et al. Cloning and sequencing of the cellular-viral junctions from the human adenovirus type 5 transformed 293 cell line. Virology. 1997. 233 (2): 423-9.