中國(guó)醫(yī)科大學(xué)附屬盛京醫(yī)院劉揚(yáng)和孫思予教授研究團(tuán)隊(duì)通過(guò)研究表明脂滴形成在肝細(xì)胞癌(HCC)細(xì)胞對(duì)索拉非尼(sorafenib)的適應(yīng)性中至關(guān)重要,并利用基于質(zhì)譜的脂質(zhì)組和蛋白質(zhì)組揭示了其中重要的調(diào)控基因和作用機(jī)制。研究成果以“AKR1C3-dependent lipid droplet formation confers hepatocellular carcinoma cell adaptability to targeted therapy”為題,發(fā)表在Theranostics(IF:11.600)上。

研究結(jié)果
1.索拉非尼抗性的肝癌細(xì)胞中存在著脂肪酸氧化(FAO)向糖酵解轉(zhuǎn)變導(dǎo)致的脂滴積累
研究者首先構(gòu)建了兩株索拉非尼抗性的肝細(xì)胞癌(HCC)細(xì)胞系,分別為HepG2R 和HuH7R。隨后,檢測(cè)了索拉非尼抗性癌細(xì)胞的耗氧率(OCR)和細(xì)胞外酸化率(ECAR)。結(jié)果顯示,抗性癌細(xì)胞的OCR下降,ECAR上升,ECAR/OCR的比率也顯著上升。糖酵解代謝流檢測(cè)發(fā)現(xiàn),糖酵解增強(qiáng)。此外,研究還發(fā)現(xiàn),抗性癌細(xì)胞完全廢棄了脂肪酸氧化(FAO)。研究者進(jìn)一步檢測(cè)了細(xì)胞內(nèi)無(wú)法被線粒體正常代謝的脂肪酸是否儲(chǔ)存在脂滴中。結(jié)果顯示,在索拉非尼和瑞戈非尼(Regorafenib)存在時(shí),HepG2R 和HuH7R細(xì)胞里均檢測(cè)到了清晰的脂滴。索拉非尼處理能增加原始癌細(xì)胞中線粒體活性氧(ROS)的產(chǎn)生,而在抗性癌細(xì)胞中僅有少量ROS的產(chǎn)生。此外,在抗性癌細(xì)胞的線粒體周?chē)鷻z測(cè)到了明顯的脂滴積累,這表明脂肪酸(FA)轉(zhuǎn)化為了甘油三酯(TAG)進(jìn)行存儲(chǔ)。為了確定脂滴形成是否對(duì)索拉非尼抗性有關(guān)鍵作用,研究者這使用Triacsin C對(duì)酰基輔酶A合成酶進(jìn)行抑制進(jìn)而阻止脂滴形成,結(jié)果抑制了索拉非尼抗性癌細(xì)胞的生長(zhǎng)。
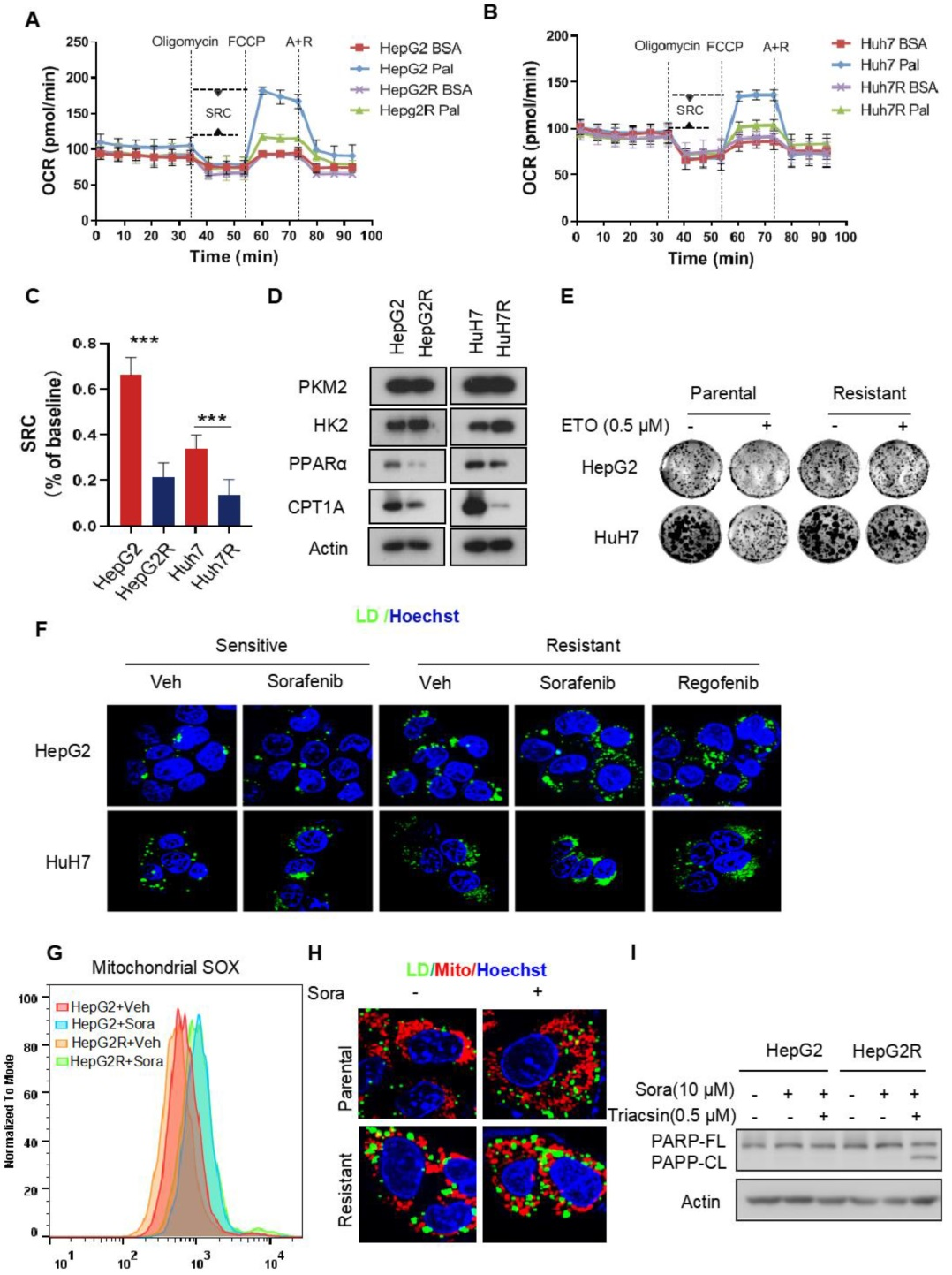
2. 脂質(zhì)組學(xué)和蛋白質(zhì)組學(xué)聯(lián)合分析發(fā)現(xiàn)AKR1C3是TAG聚集的調(diào)控因子
接下來(lái),研究者對(duì)抗性癌細(xì)胞和原始癌細(xì)胞進(jìn)行了脂質(zhì)組學(xué)檢測(cè),結(jié)果顯示TAG(甘油三酯)是在抗性癌細(xì)胞中上調(diào)最顯著的代謝物,此外,前列腺素(PG)在抗性癌細(xì)胞中也普遍上調(diào)。為了找到調(diào)控TAG聚集的關(guān)鍵蛋白質(zhì),研究者對(duì)抗性癌細(xì)胞和原始癌細(xì)胞分別進(jìn)行了蛋白質(zhì)組學(xué)檢測(cè),有5個(gè)蛋白,包括HSD17B5 (AKR1C3)、TYB10、MGST1、 FSCN1和ANXA2在抗性癌細(xì)胞中上調(diào)。已有報(bào)道表明,AKR1C3與多囊卵巢綜合征(PCOS)脂肪細(xì)胞中的脂質(zhì)代謝和脂質(zhì)形成相關(guān),因此,研究者決定探究AKR1C3在肝細(xì)胞癌脂質(zhì)代謝中的作用。
WB證實(shí)了AKR1C3在抗性癌細(xì)胞中的高表達(dá)。在抗性癌細(xì)胞中還檢測(cè)到了脂肪酸合成酶(FASN)表達(dá)和乙酰輔酶A羧化酶(ACC)活性的上升。研究者構(gòu)建了AKR1C3過(guò)表達(dá)的原始癌細(xì)胞HepG2-AKR1C3,并進(jìn)行了脂質(zhì)組學(xué)檢測(cè),結(jié)果顯示HepG2-AKR1C3的脂質(zhì)組學(xué)特征與抗性癌細(xì)胞的脂質(zhì)組學(xué)特性相似,都有上升的甘油三酯(TAG)和前列腺素(PG)。此外,AKR1C3過(guò)表達(dá)通過(guò)下調(diào)p-ACC-s79和上調(diào)FASN的表達(dá)促進(jìn)了脂質(zhì)生成和脂滴產(chǎn)生,這與抗性癌細(xì)胞HepG2R中觀察到的現(xiàn)象一致。綜合以上結(jié)果表明,AKR1C3對(duì)脂滴形成有關(guān)鍵作用。
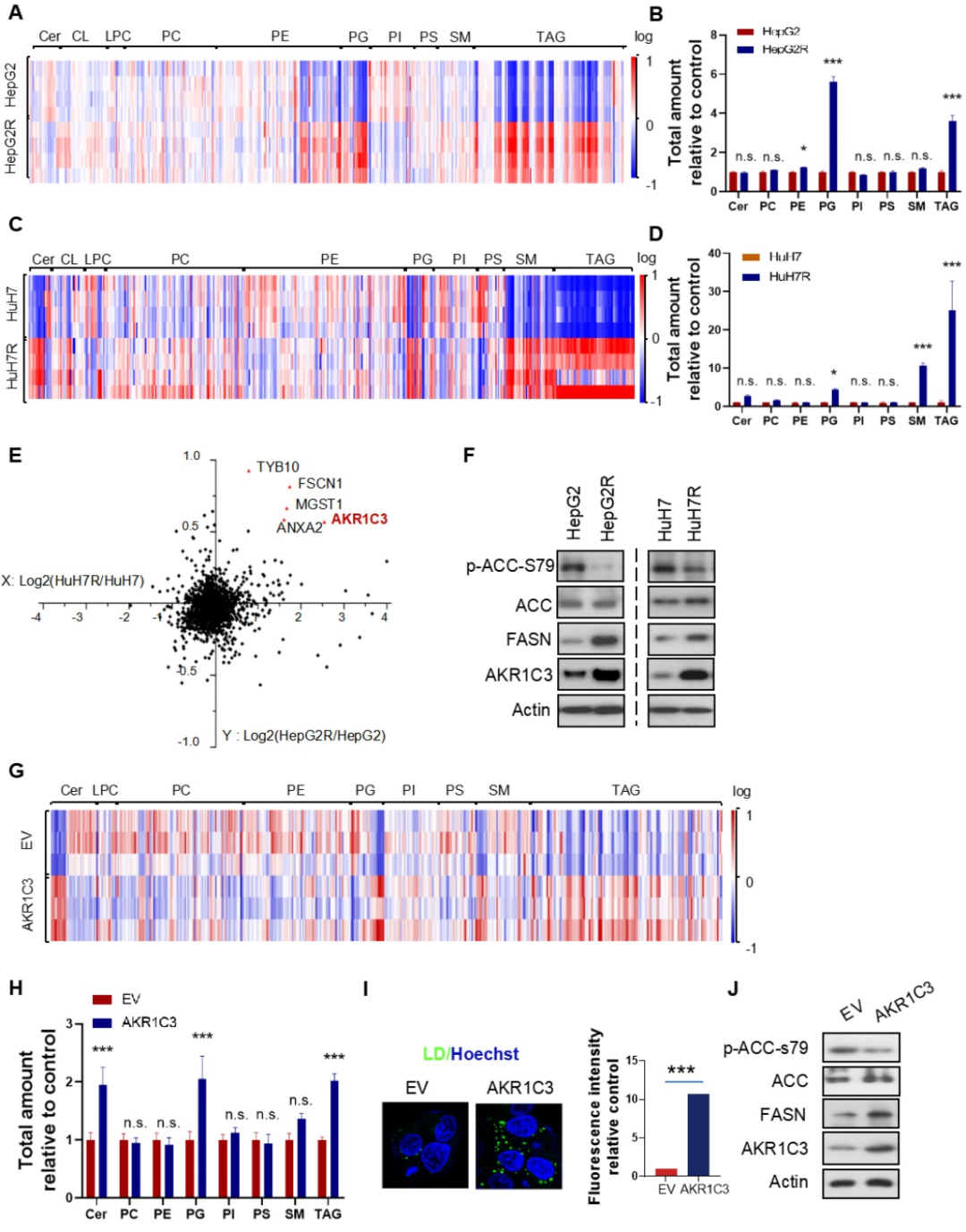
3. AKR1C3的缺失廢除TAG(甘油三酯)的積累并刺激FAO(脂肪酸氧化)
為了確定AKR1C3缺失是否影響肝細(xì)胞癌的脂質(zhì)積累,研究者利用CRISPR/Cas9系統(tǒng)敲除了抗性癌細(xì)胞HepG2R中的AKR1C3并進(jìn)行脂質(zhì)組學(xué)檢測(cè)。結(jié)果表明,最明顯改變的代謝物富集在甘油磷脂代謝、甘油脂代謝和糖基磷脂酰肌醇(GPI)-錨定蛋白生物合成途徑。此外,AKR1C3的缺失明顯導(dǎo)致甘油三酯(TAG)的耗竭,以及二酰基甘油(DAG)、甘油磷脂(PC、LPC、PI、PS)和鞘磷脂(Cer、SM)種類(lèi)的上調(diào)。此外,與對(duì)照組相比,AKR1C3-/-HepG2R細(xì)胞中的線粒體β-氧化增強(qiáng)。在AKR1C3-/-細(xì)胞中也明顯觀察到線粒體膜電位和活性氧(ROS)生成的增加。總之,這些結(jié)果證明,AKR1C3的缺失抑制了細(xì)胞內(nèi)甘油三酯(TAG)的積累,促進(jìn)了脂肪酸運(yùn)輸?shù)骄€粒體進(jìn)行β-氧化。
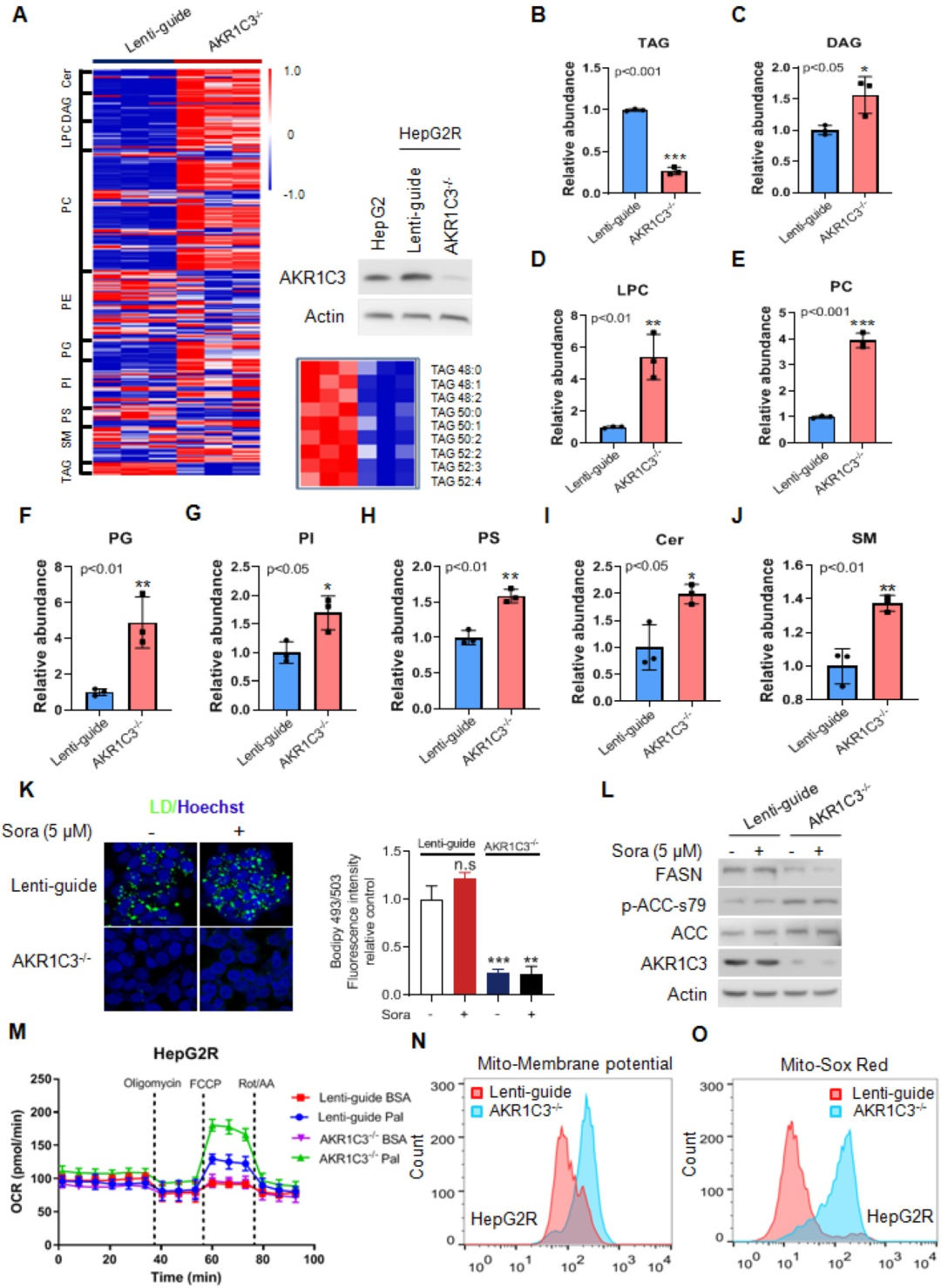
4. AKR1C3依賴的脂滴形成阻止線粒體功能障礙和脂肪毒性
研究者假設(shè)AKR1C3通過(guò)抑制脂滴分解促進(jìn)脂滴形成。因?yàn)樵贏KR1C3-/-細(xì)胞中存在脂滴積累,于是研究者使用兩種AKR1C3的抑制劑,檢測(cè)當(dāng)AKR1C3活性被抑制后,是否會(huì)引起脂滴分解。研究者對(duì)AKR1C3抑制劑FLU(氟芬那酸)或5-βCA(5β-膽烷酸)處理后的抗性癌細(xì)胞HepG2R分別進(jìn)行脂質(zhì)組學(xué)檢測(cè)。結(jié)果與AKR1C3-/-細(xì)胞相一致,在索拉非尼存在的條件下,抑制AKR1C3顯著降低細(xì)胞甘油三酯(TAG)和脂滴的含量,而磷脂和鞘脂類(lèi)脂質(zhì)包括PE、PG、PI、PS和SM的含量則上升。同時(shí)還檢測(cè)到抑制AKR1C3能顯著降低甘油三酯(TAG)的減少以及細(xì)胞內(nèi)游離脂肪酸(FFA)的增加,提示線粒體中β-氧化作用的減弱。研究者對(duì)線粒體功能的檢測(cè)也發(fā)現(xiàn)抑制AKR1C3造成線粒體功能障礙。
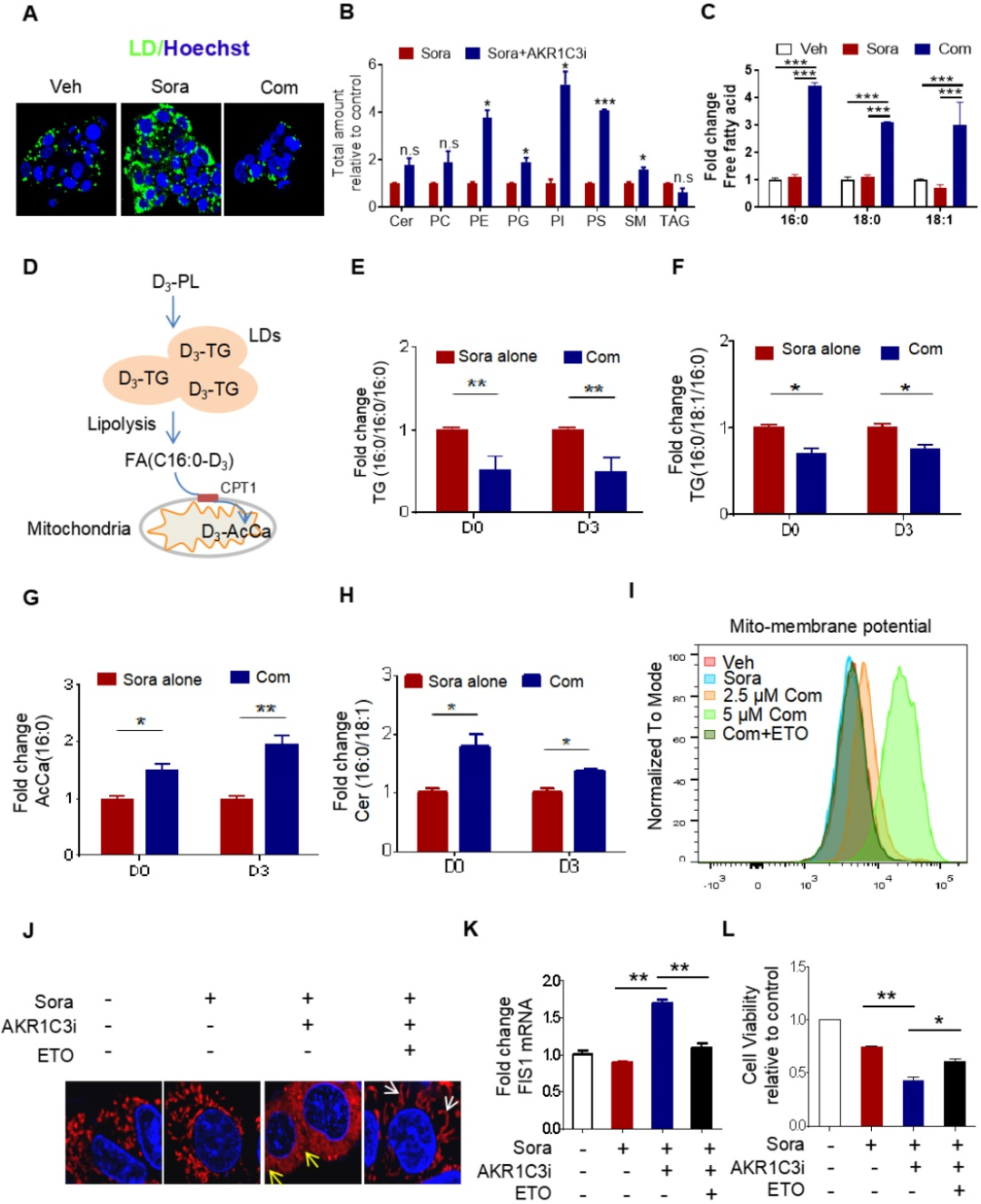
5. AKR1C3抑制脂質(zhì)自噬以支持脂滴形成
接下來(lái)研究者探究AKR1C3是如何調(diào)控脂質(zhì)積累的。鑒于AKR1C3的基本功能是調(diào)控PG(前列腺素)合成,研究者首先檢測(cè)了PG代謝是如何促進(jìn)脂質(zhì)積累的。結(jié)果表明AKR1C3介導(dǎo)的索拉非尼抗性和脂滴積累是PG非依賴性的。
已有研究表明,抑制AKR1C3能增強(qiáng)胃腸癌細(xì)胞系中阿霉素誘導(dǎo)的細(xì)胞自噬,因此研究者假設(shè)AKR1C3可能通過(guò)自噬介導(dǎo)的脂滴分解(脂質(zhì)自噬)調(diào)控脂滴穩(wěn)態(tài)。通過(guò)mCherry-GFP-LC3 融合蛋白實(shí)驗(yàn)表明,抑制AKR1C3能增強(qiáng)細(xì)胞自噬。加強(qiáng)細(xì)胞自噬能在AKR1C3抑制的條件下誘導(dǎo)更顯著的脂滴清除,而這種清除作用會(huì)被自噬抑制劑減弱。進(jìn)一步研究發(fā)現(xiàn),在AKR1C3抑制劑和索拉非尼聯(lián)合處理?xiàng)l件下,自噬/脂質(zhì)自噬的抑制或者敲低ATG5都能顯著增強(qiáng)細(xì)胞生存,減少凋亡細(xì)胞數(shù)量,細(xì)胞顯示出減少的細(xì)胞色素C釋放和活性氧(ROS)生成,并有脂滴的增加,且線粒體膜電位能恢復(fù)到正常區(qū)間。綜合以上結(jié)果表明,AKR1C3通過(guò)脂質(zhì)自噬調(diào)控脂滴穩(wěn)態(tài)。
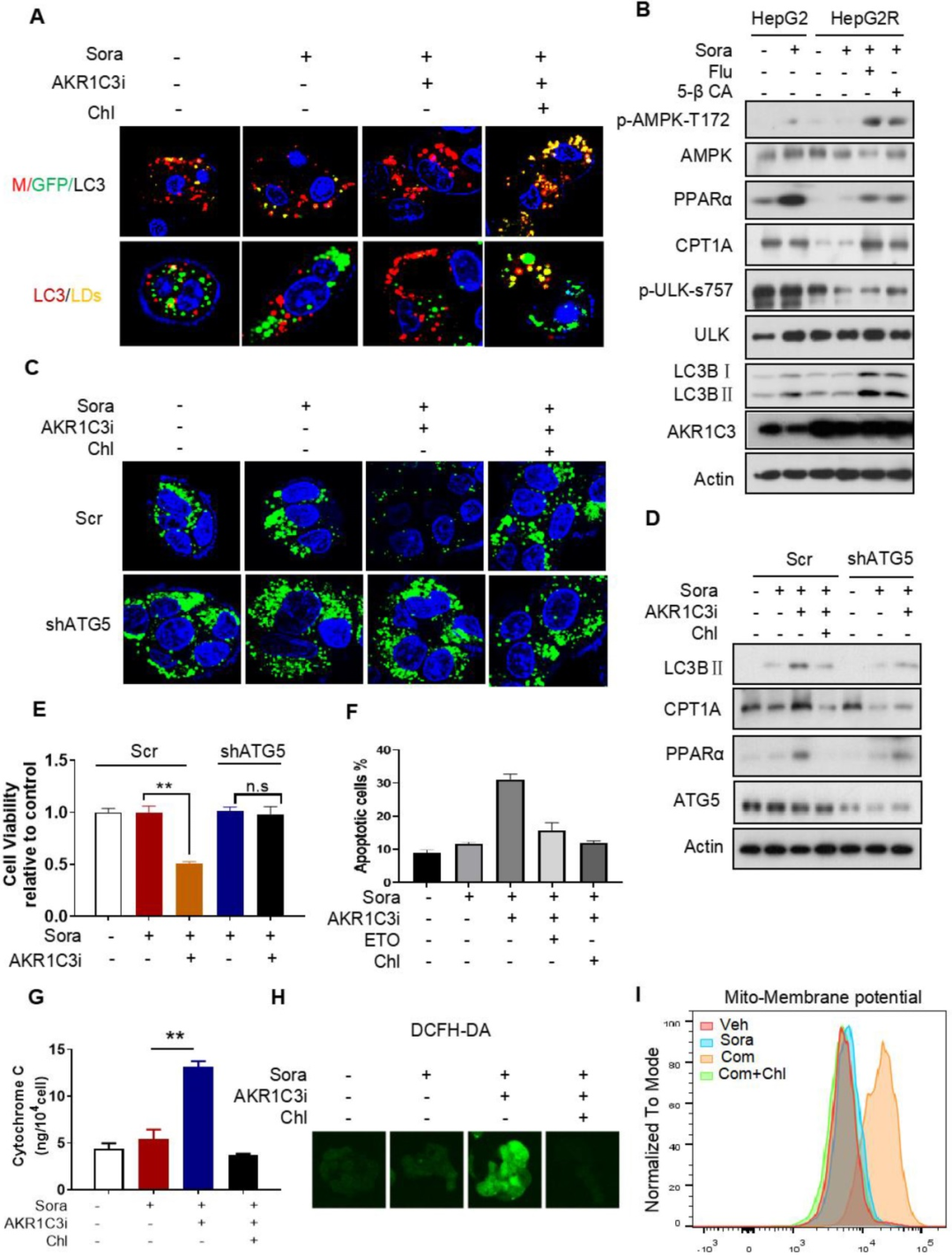
6. AKR1C3促進(jìn)代謝從氧化磷酸化轉(zhuǎn)向糖酵解
為了檢測(cè)AKR1C3是否會(huì)影響能量代謝,研究者對(duì)原始癌細(xì)胞中過(guò)表達(dá)AKR1C3或在抗性癌細(xì)胞中敲低AKR1C3。過(guò)表達(dá)AKR1C3后,原始癌細(xì)胞的耗氧率(OCR)下降,細(xì)胞外酸化率(ECAR)上升,ECAR/OCR比值上調(diào)3.7倍。在抗性癌細(xì)胞中敲低AKR1C3后,耗氧率(OCR)上升,細(xì)胞外酸化率(ECAR)下降,,ECAR/OCR比值下降2倍。此外,過(guò)表達(dá)AKR1C3降低了AMPK-T172磷酸化、PPARα和CPT1A的水平,但增加了HK2的蛋白水平。而AKR1C3的抑制恢復(fù)了索拉非尼處理的癌細(xì)胞中上述蛋白的水平。此外,與對(duì)照細(xì)胞相比,高表達(dá)AKR1C3的HepG2細(xì)胞對(duì)葡萄糖表現(xiàn)出更大的生長(zhǎng)依賴性。相反,與對(duì)照組相比,AKR1C3-/- HepG2R細(xì)胞對(duì)葡萄糖的敏感性較低。最后,在索拉非尼處理?xiàng)l件下,AKR1C3過(guò)表達(dá)明顯提高了HepG2細(xì)胞的存活率,而敲除AKR1C3使HepG2R細(xì)胞對(duì)索拉非尼敏感。這些結(jié)果表明,AKR1C3使代謝從脂肪酸氧化轉(zhuǎn)向糖酵解,以促進(jìn)HCC細(xì)胞對(duì)索拉非尼的抗性。
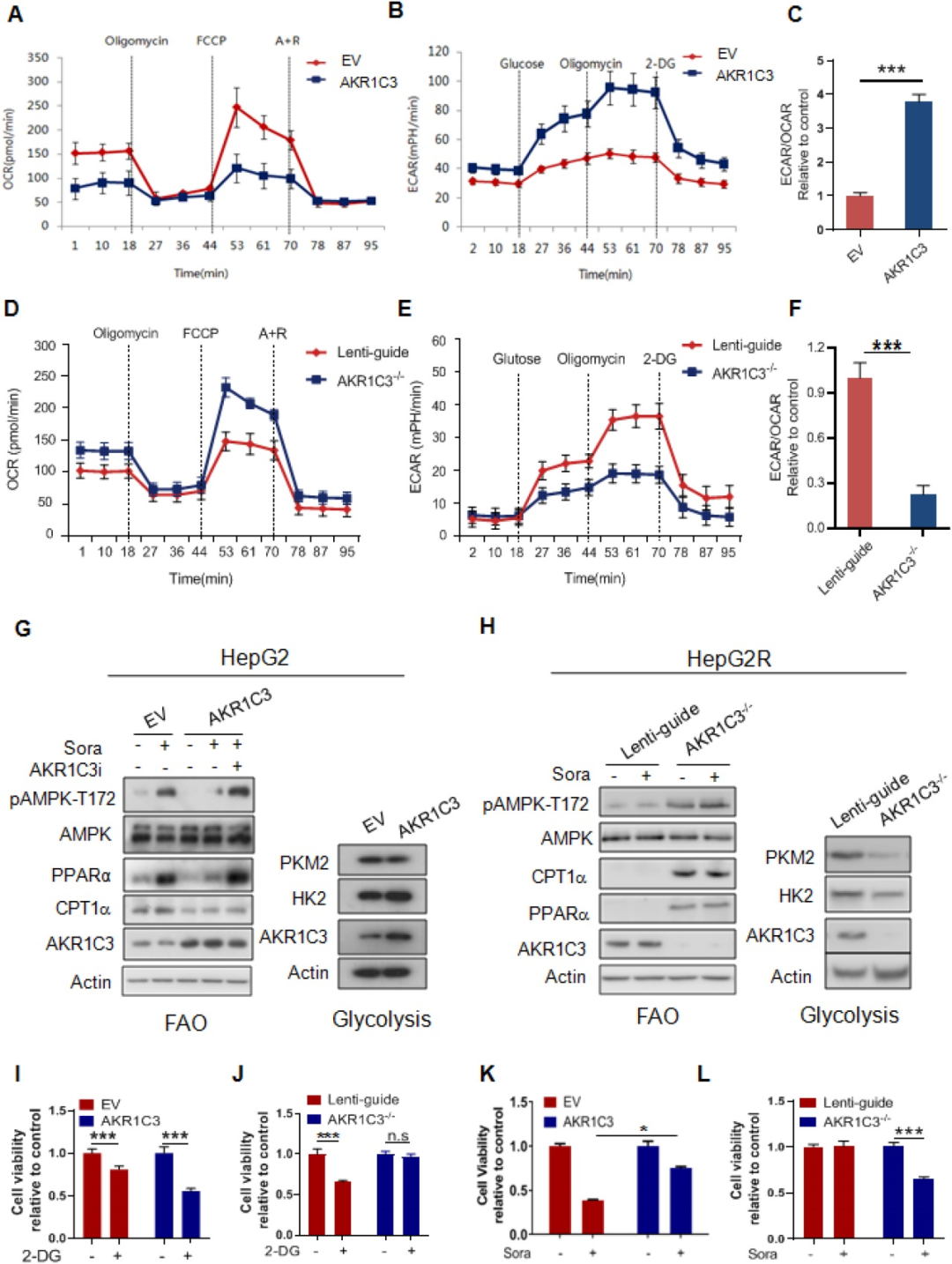
7. AKR1C3維持索拉非尼處理?xiàng)l件下肝細(xì)胞癌細(xì)胞的生長(zhǎng)
接下來(lái),研究者探究了索拉非尼和FLU (氟芬那酸,AKR1C3抑制劑)聯(lián)合治療對(duì)肝細(xì)胞癌細(xì)胞的影響。結(jié)果表明,聯(lián)合治療強(qiáng)烈地抑制了抗性癌細(xì)胞HepG2R和HuH7R異種移植瘤的生長(zhǎng)。免疫組化顯示CPT1A和PPARα的表達(dá)明顯升高,提示細(xì)胞存在脂滴分解。油紅染色實(shí)驗(yàn)證明,聯(lián)合治療降低了肝細(xì)胞癌細(xì)胞異種移植腫瘤中的脂滴積累。進(jìn)一步,研究者觀察了AKR1C3-/-HepG2R移植瘤小鼠的腫瘤生長(zhǎng)情況。結(jié)果表明,AKR1C3缺失與索拉非尼協(xié)同作用,通過(guò)誘導(dǎo)脂肪吞噬來(lái)抑制腫瘤生長(zhǎng)。以上基于肝細(xì)胞癌細(xì)胞異種移植瘤模型的研究表明,抑制AKR1C3會(huì)誘導(dǎo)脂滴分解和線粒體脂質(zhì)毒性,從而使耐藥的肝細(xì)胞癌細(xì)胞對(duì)索拉非尼敏感。
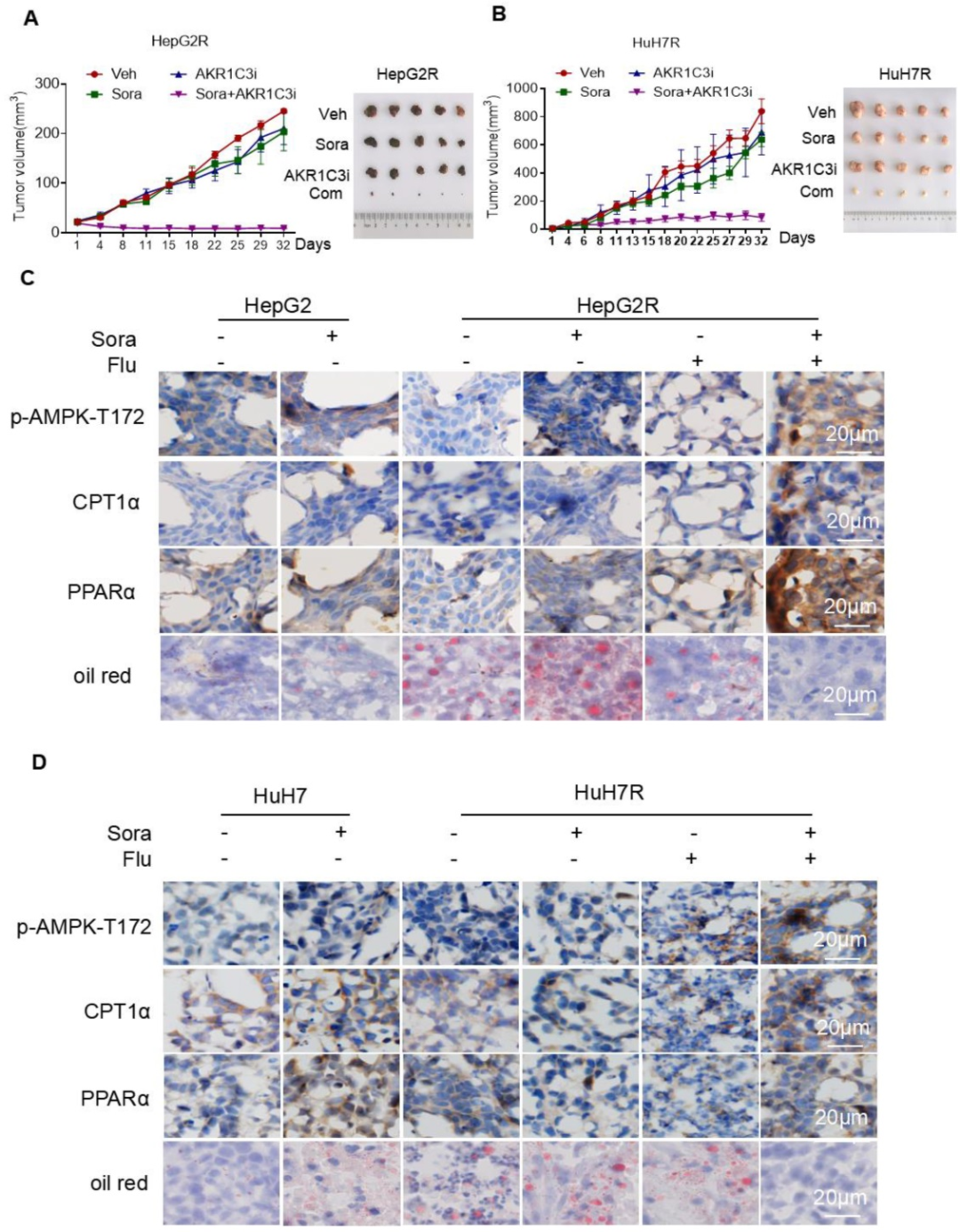
研究總結(jié)
本研究發(fā)現(xiàn),長(zhǎng)期的索拉非尼治療會(huì)損害脂肪酸氧化(FAO),導(dǎo)致肝細(xì)胞癌(HCC)細(xì)胞中脂滴(LD)的積累。對(duì)索拉非尼抗性和原始癌細(xì)胞的脂質(zhì)組學(xué)和蛋白質(zhì)組學(xué)聯(lián)合分析發(fā)現(xiàn),醛酮還原酶AKR1C3是肝細(xì)胞癌中脂滴積累的重要調(diào)控基因。AKR1C3缺失完全消除脂滴含量,使游離脂肪酸流向磷脂、鞘脂和線粒體。此外,研究者發(fā)現(xiàn)AKR1C3依賴的脂滴積累是緩解索拉非尼誘導(dǎo)的線粒體脂肪毒性和功能障礙的必要條件。藥物抑制AKR1C3的活性會(huì)立即誘導(dǎo)自噬依賴的脂滴分解,導(dǎo)致線粒體裂變和索拉非尼抗性肝細(xì)胞癌細(xì)胞凋亡。改變AKR1C3的表達(dá)足以驅(qū)動(dòng)脂肪酸氧化和糖酵解之間的代謝轉(zhuǎn)換。AKR1C3可作為肝細(xì)胞癌治療的一個(gè)潛在靶標(biāo)。
吉?jiǎng)P基因憑借多年在靶標(biāo)篩選及驗(yàn)證服務(wù)領(lǐng)域的技術(shù)積累,建立的標(biāo)準(zhǔn)化 、工程化 、系統(tǒng)化的GRP平臺(tái),為中國(guó)研究型醫(yī)生提供科研服務(wù),加快科研成果轉(zhuǎn)化。其中,多組學(xué)平臺(tái)包含蛋白質(zhì)組學(xué)平臺(tái)和高通量測(cè)序平臺(tái):
·蛋白質(zhì)組學(xué)平臺(tái)擁有多臺(tái)timsTOF Pro、Exploris 480高精度質(zhì)譜儀,專(zhuān)業(yè)的Spectronaut Plusar、Mascot等分析軟件,提供專(zhuān)業(yè)的4D、DIA、TMT、PRM、磷酸化修飾組、olink蛋白質(zhì)組等檢測(cè)服務(wù),強(qiáng)大的機(jī)器學(xué)習(xí)算法、IPA分析、蛋白基因組分析服務(wù),系統(tǒng)的生物標(biāo)志物、分子分型、藥物靶點(diǎn)、基因功能研究等解決方案,真正讓廣大研究型醫(yī)生的科研工作更省心、更省力、更高效;
·高通量測(cè)序平臺(tái)分為常規(guī)測(cè)序服務(wù)和單細(xì)胞測(cè)序服務(wù):單細(xì)胞測(cè)序擁有10x和BD兩個(gè)平臺(tái),提供單細(xì)胞RNA-seq、單細(xì)胞核測(cè)序、單細(xì)胞混樣RNA-seq、單細(xì)胞TCR/BCR、單細(xì)胞(RNA+ATAC)、空間轉(zhuǎn)錄組測(cè)序等服務(wù);常規(guī)測(cè)序服務(wù)提供meRIP-seq(m6A/m1A/m7G/m5C 等RNA甲基化修飾測(cè)序)、acRIP-seq(ac4C RNA乙酰化修飾測(cè)序)、ATAC-seq、Ribo-seq(翻譯組測(cè)序) 、mRNA/miRNA/LncRNA/circRNA-seq、全轉(zhuǎn)錄組測(cè)序(兩文庫(kù)/三文庫(kù))、外泌體miRNA/LncRNA-seq、WGS/WES、WGBS、RRBS、BSAS等服務(wù)。