確保外源基因在細(xì)胞或者動物體內(nèi),持續(xù)和高水平的表達(dá)具有廣泛的應(yīng)用需求及科研價(jià)值。當(dāng)前遞送外源基因的生物學(xué)手段,主要是運(yùn)用質(zhì)粒或者病毒作為媒介表達(dá)基因,而啟動子是實(shí)現(xiàn)基因表達(dá)的基礎(chǔ)元件。但是,經(jīng)常有老師反饋,為什么基因表達(dá)不強(qiáng)呢?為什么細(xì)胞傳代幾次之后就不表達(dá)了呢?今天,小編帶您盤點(diǎn)下,常用的組成型啟動子表達(dá)效率與細(xì)胞的關(guān)系。
啟動子是位于結(jié)構(gòu)基因上游的一段可以行使轉(zhuǎn)錄起始功能的序列,結(jié)構(gòu)如下:
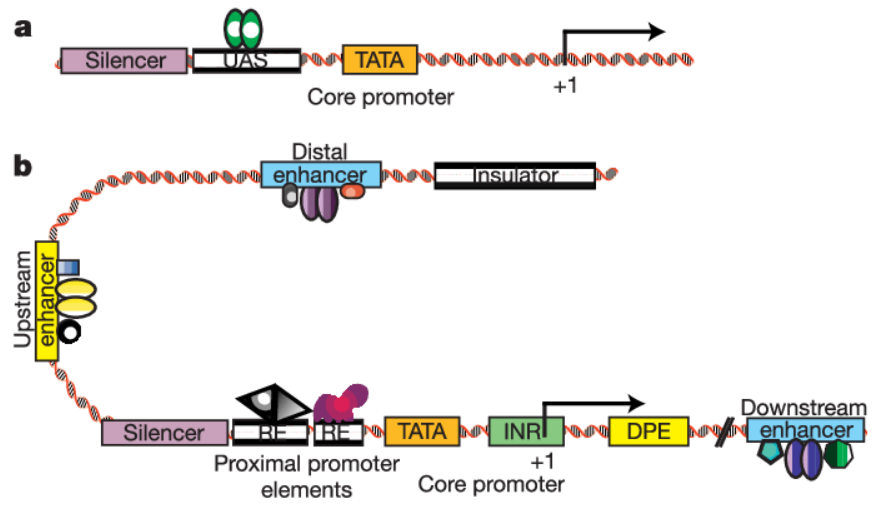
根據(jù)其對α-鵝膏蕈堿的敏感性不同,分為三類,常用的啟動子是Ⅱ、Ⅲ兩種啟動子。其中,Ⅲ型啟動子主要表達(dá)非編碼的短序列RNA,比如shRNA、miRNA等,我們常說的啟動子,或者細(xì)胞里面常規(guī)基因的啟動子,幾乎都是Ⅱ型啟動子,其特點(diǎn)是轉(zhuǎn)錄產(chǎn)物長度不限,編碼與否不限等特點(diǎn)。
酶的種類1 | 存在 | 功能 | 對抑制物的敏感性 |
RNA聚合酶I | 核仁 | 合成rRNA前體 | 不敏感 |
RNA聚合酶II | 核質(zhì) | 合成mRNA前體及大多數(shù)snRNA | 敏感 |
RNA聚合酶III | 核質(zhì) | 合成5S rRNA前體、tRNA前體及其他的核和胞質(zhì)小RNA前體 | 存在物種特異性 |
Ⅱ型啟動子在使用特點(diǎn)上又可分為三大類:組成型啟動子、特異性啟動子、誘導(dǎo)啟動子。特異性啟動子只能在特定細(xì)胞表達(dá),因此適合動物實(shí)驗(yàn)使用;誘導(dǎo)啟動子需要加入特定的藥物才能激發(fā)或者抑制其活性,如四環(huán)素誘導(dǎo)TRE表達(dá);而表達(dá)系統(tǒng)中,運(yùn)用最多的則是組成型啟動子,這類啟動子在絕大部分細(xì)胞里面都能維持一定的表達(dá)活性,當(dāng)然,不同的啟動子也有高低表達(dá)之分。
載體上最常見的組成型啟動子包含:β-肌動蛋白啟動子(ACTB),巨細(xì)胞病毒(CMV),延伸因子-1α(EF1α),磷酸甘油酸激酶(PGK)、泛素C(UbC、本文別名FCIV),具有CMV最大內(nèi)含子的CMV啟動子/增強(qiáng)子(CMVi啟動子)、巨細(xì)胞病毒增強(qiáng)子和雞β-肌動蛋白啟動子組成的CAG,猿猴病毒來源的SV40等。其中,最常見的是CMV啟動子,在大量的商業(yè)化載體上如pcDNA3、pEGFP使用,病毒上最常見啟動子是CMV、EF1α,說明這些都是高表達(dá)啟動子,那么所有細(xì)胞系都合適嗎?小編收羅了不同文獻(xiàn)、不同細(xì)胞類型的驗(yàn)證實(shí)驗(yàn),接著往下看:
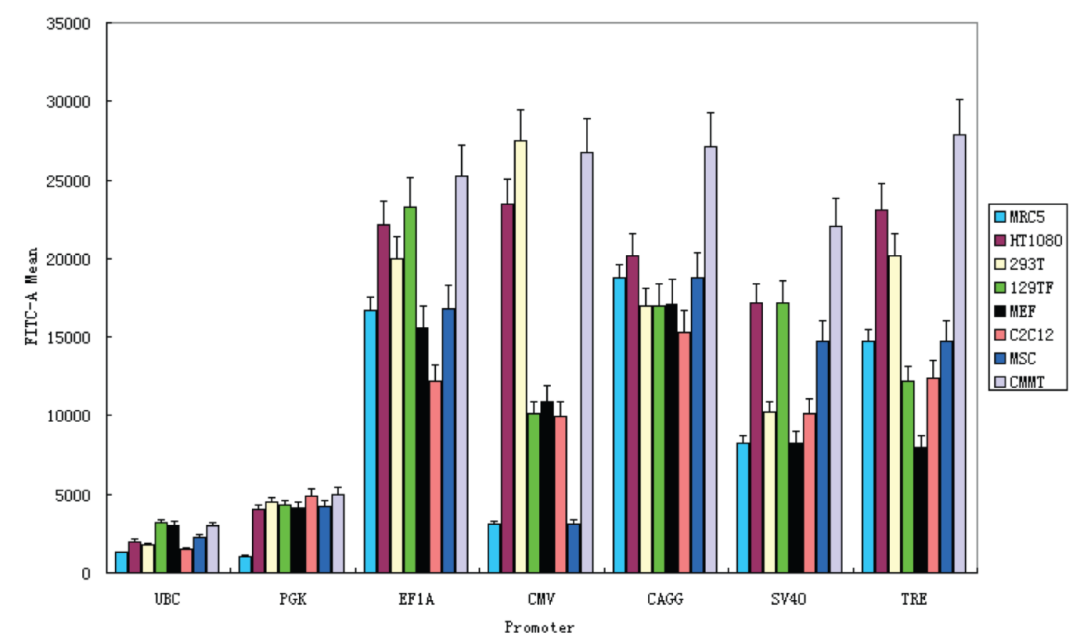
混合細(xì)胞系
作者選了多種來源的細(xì)胞,包含成纖維細(xì)胞、腫瘤細(xì)胞、干細(xì)胞、高度分化的肌肉細(xì)胞等,使用六個哺乳動物組成型啟動子,每個都插入GFP包裝成慢病毒轉(zhuǎn)導(dǎo)細(xì)胞,通過流式細(xì)胞儀對GFP強(qiáng)度進(jìn)行定量:
小鼠尾巴成纖維細(xì)胞(129TF),小鼠胚胎成纖維細(xì)胞(MEF),小鼠成肌細(xì)胞(C2C12),大鼠間充質(zhì)干細(xì)胞(MSC),人成纖維細(xì)胞(MRC5),人纖維肉瘤細(xì)胞(HT1080),人胚胎腎細(xì)胞(293T)和恒河猴獼猴乳腺腫瘤細(xì)胞(CMMT)
結(jié)果表明:UBC跟PGK始終是弱的,SV40相對較強(qiáng),EF1A和CAG啟動子在所有細(xì)胞類型中均具有較強(qiáng)的一致性,CMV啟動子是最多變的,是在一些細(xì)胞類型(很強(qiáng)烈例如,293T和CMMT)和其他(較弱例如,MRC5和MSC)。
由此看出,CMV在腫瘤細(xì)胞系傾向高表達(dá),而EF1A和CAG在大部分細(xì)胞表達(dá)趨于穩(wěn)定,且這三個高強(qiáng)度啟動子在干細(xì)胞等類型活性都有所下降。所以,針對常規(guī)的腫瘤細(xì)胞系,常規(guī)體細(xì)胞系,三個啟動子的效果還是不錯的,但由干細(xì)胞表達(dá)效果差,推導(dǎo)出非常規(guī)的細(xì)胞系,如懸浮細(xì)胞系、神經(jīng)細(xì)胞等,啟動子如何選擇呢?
胚胎干細(xì)胞(hESCs)
干細(xì)胞相對復(fù)雜,因?yàn)檫€存在分化過程的干細(xì)胞,那么干細(xì)胞對于啟動子的選擇有何特點(diǎn)呢?
對hESCs中ACTB,CMV,EF1α,PGK和UbC啟動子的活性進(jìn)行了比較研究。選擇慢病毒介導(dǎo)的基因轉(zhuǎn)移作為基因傳遞系統(tǒng),通過FACS分選出了eGFP +細(xì)胞,結(jié)果如下:
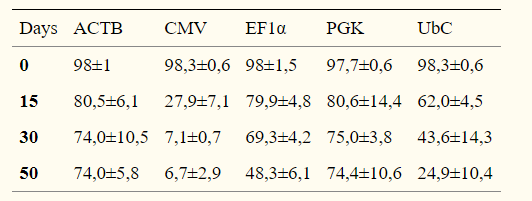
1.未分化的hESCs的啟動子活性
在FACS分選后的15、30和50天測量啟動子活性,通過OCT3 / 4,NANOG和hES-Cellect(Cellartis AB)多能蛋白的標(biāo)記證實(shí)了eGFP +細(xì)胞的多能性:
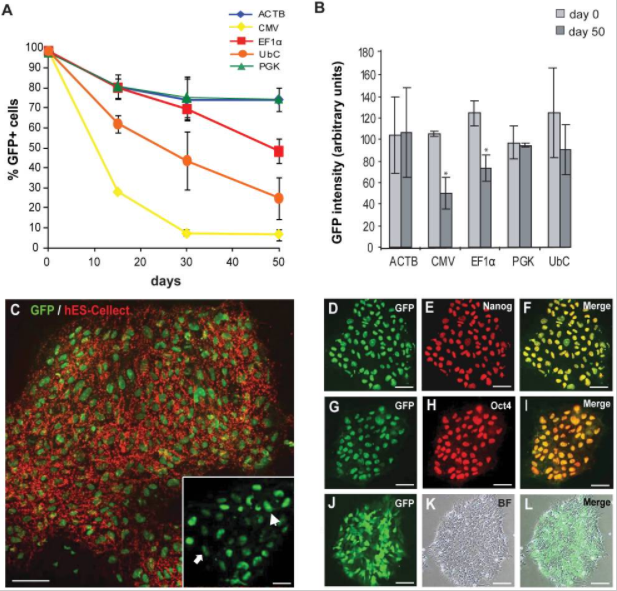
第0天,所有啟動子的eGFP+百分比約為98%,第30天,ACTB,EF1α和PGK的啟動子活性均相等,第50天,CMV-eGFP+細(xì)胞的百分比降低于10%,EF1α也出現(xiàn)下降。
2. 分化的hESCs中的啟動子活性
慢病毒介導(dǎo)GFP的將hESC系SA121,在第0天(d0),未分化狀態(tài)第22天(undiff),分化為胚狀體22天(EB),測試GFP +細(xì)胞的比例,結(jié)果顯示,啟動子在分化過程中被顯著下調(diào),并且在大約50%的分化細(xì)胞中沒有活性。EF1α是分化過程中最穩(wěn)定的啟動子;CMV啟動子僅在細(xì)胞的一小部分(約15%)具有活性。
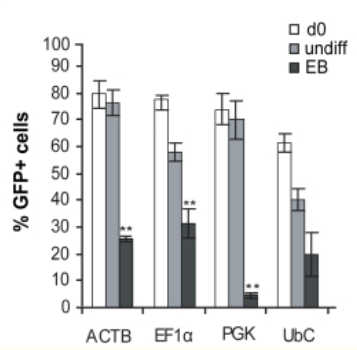
總之,就長期保持未分化的hESCs的轉(zhuǎn)基因(eGFP)表達(dá)而言,ACTB,EF1α和PGK啟動子是最穩(wěn)定的啟動子;而在分化過程中,成型啟動子的活性會下降,相對而言,EF1α啟動子具有最高的穩(wěn)定性。原因即細(xì)胞在分化過程中,所表達(dá)的轉(zhuǎn)錄因子是有差異的,部分啟動子表達(dá)所需的轉(zhuǎn)錄因子丟失,同時(shí)染色體結(jié)構(gòu)的改變影響了外源插入序列的狀態(tài),導(dǎo)致啟動子出現(xiàn)沉默現(xiàn)象。
造血干細(xì)胞(HSC)/祖細(xì)胞
造血干細(xì)胞(HSC)具有自我更新和多譜系分化成所有成熟血細(xì)胞的潛力。因此,有效轉(zhuǎn)導(dǎo)至HSCs將提供治療因血細(xì)胞功能異常而導(dǎo)致的多種疾病的機(jī)會,并且將成為研究HSCs增殖,分化和運(yùn)輸調(diào)節(jié)的有力工具。作者構(gòu)建了攜帶多個啟動子表達(dá)熒光的腺病毒,感染兩種狀態(tài)的造血細(xì)胞,以流式檢測:
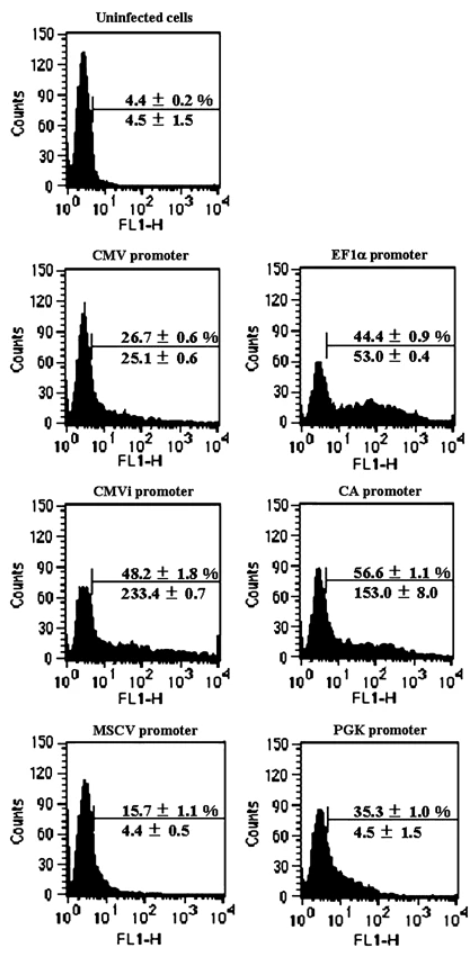
成熟CD34+ 成熟CD34+
免疫細(xì)胞
結(jié)果顯示:當(dāng)用于造血細(xì)胞中時(shí),CMV啟動子傾向于沉默。轉(zhuǎn)基因可能以非常低的水平表達(dá)或無法檢測。EF1α、CMVi和CAG啟動子在轉(zhuǎn)導(dǎo)人骨髓CD34+細(xì)胞方面表現(xiàn)優(yōu)異。特別地,CAG啟動子在CD34+細(xì)胞和未成熟的CD34+亞群中最有效地發(fā)揮作用。
當(dāng)前,以CAR-T和TCR-T為代表的細(xì)胞治療非常火熱,前者都需要運(yùn)用慢病毒感染T細(xì)胞。免疫類細(xì)胞,諸如T細(xì)胞,病毒感染難度較大,啟動子的選擇得注意。
作者構(gòu)建多個啟動子的慢病毒載體表達(dá)熒光,感染經(jīng)過IL-2刺激過的PBL細(xì)胞,以FACS檢測,觀察到CMV啟動子在原代T細(xì)胞驅(qū)動效率低,CAG也相對較差,其余幾個啟動子的效率相近,都在60%上下。
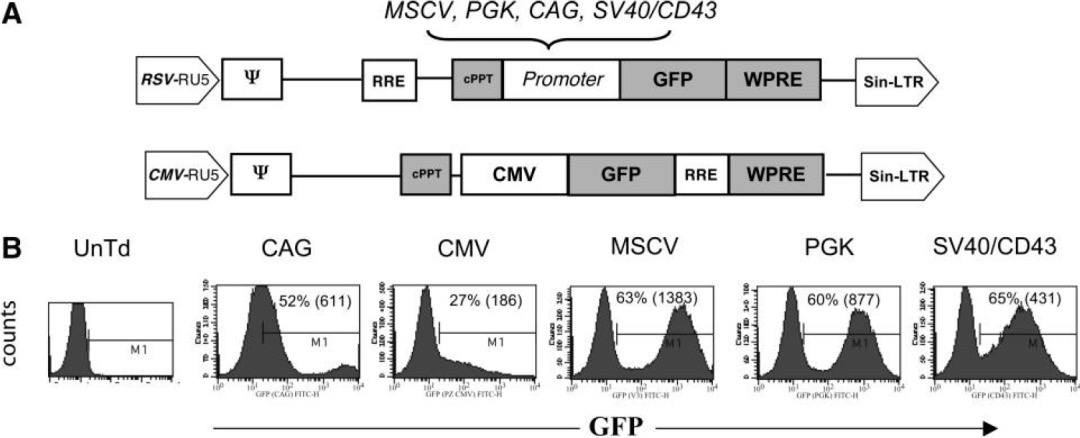
進(jìn)一步以以抗CD3抗體OKT3刺激細(xì)胞,再進(jìn)行實(shí)驗(yàn),活化后的T細(xì)胞,多個啟動子的表達(dá)效率得到增強(qiáng),其中以MSCV啟動子最優(yōu)。
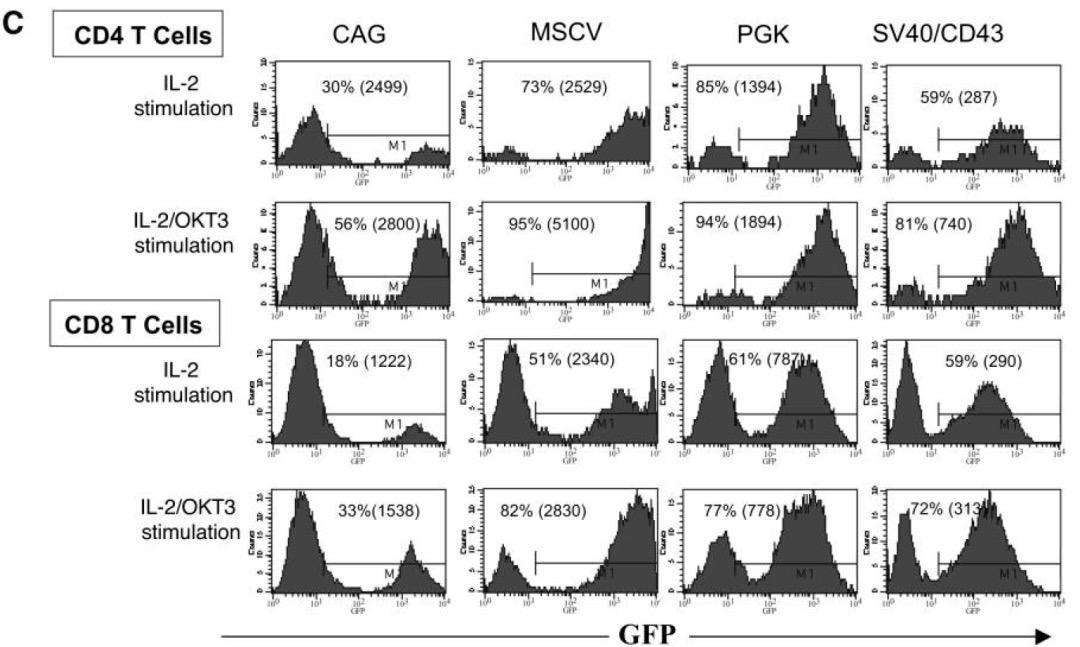
在PBL中,發(fā)現(xiàn)含有MSCV啟動子的載體最適合,在最低程度刺激的淋巴細(xì)胞和高度活化的淋巴細(xì)胞中的表達(dá)。當(dāng)然,文章沒有比較EF1α,實(shí)際后者在T細(xì)胞也有不俗的表達(dá)水平。
神經(jīng)細(xì)胞
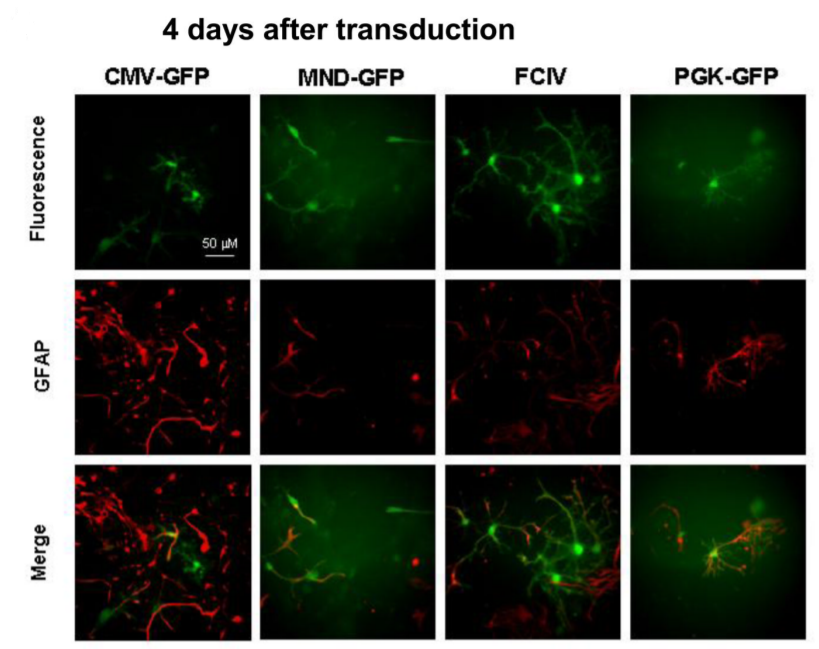
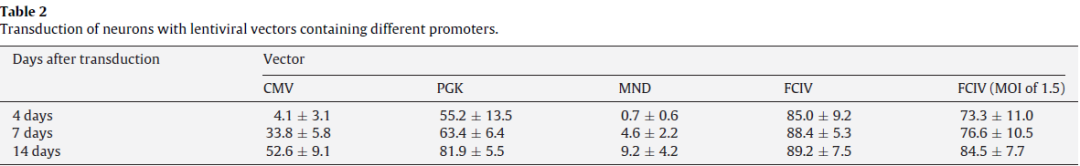
用CMV-GFP,PGK-GFP,MND-GFP和泛素C啟動子(FCIV)轉(zhuǎn)導(dǎo)了含有神經(jīng)元和星形膠質(zhì)細(xì)胞的小鼠新皮層培養(yǎng)物。轉(zhuǎn)導(dǎo)后四天,我們對神經(jīng)元(NeuN陽性)進(jìn)行檢測,在FCIV轉(zhuǎn)導(dǎo)的培養(yǎng)物中85%的神經(jīng)元表達(dá)了GFP,PGK為55.2%,且大多數(shù)報(bào)告基因陽性細(xì)胞與NeuN共定位;而CMV為4.1%、MND為0.7%。且?guī)缀跖cNeuN染色的細(xì)胞不重疊,說明后兩者不適合神經(jīng)元的表達(dá)。
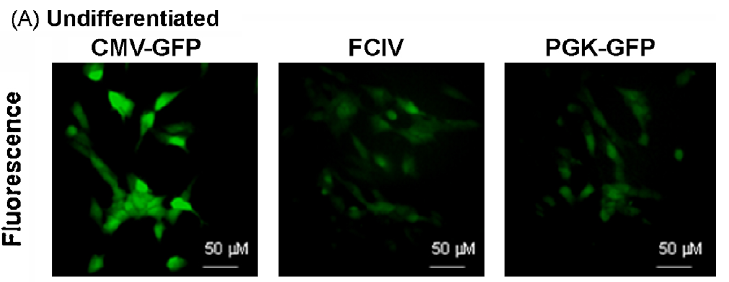
以上述病毒轉(zhuǎn)導(dǎo)來自SH-SY5Y神經(jīng)母細(xì)胞瘤細(xì)胞系,結(jié)果顯示:CMV,PGK和FCIV在未分化和分化的神經(jīng)母細(xì)胞瘤細(xì)胞中均具有活性,但CMV啟動子活性最高,尤其是在分化后(小腦顆粒神經(jīng)元顯示出相同的結(jié)果,數(shù)據(jù)見原文)。
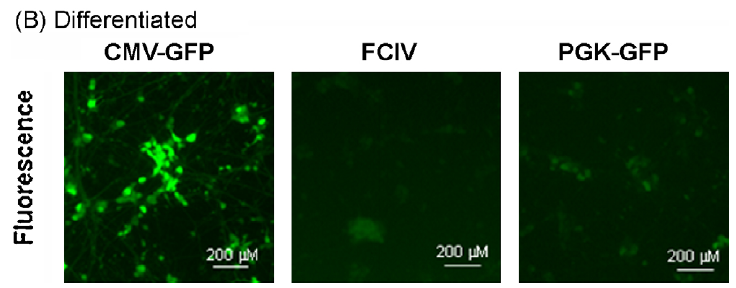
對于涉及原代皮層神經(jīng)元培養(yǎng)的研究,UBC和PGK啟動子可能是基因表達(dá)的最佳選擇。UBC,PGK,CMV和MND啟動子均適用于轉(zhuǎn)導(dǎo)原代星形膠質(zhì)細(xì)胞。在小腦顆粒細(xì)胞和分化的SH-SY5Y神經(jīng)母細(xì)胞瘤培養(yǎng)物中,CMV啟動子指導(dǎo)最穩(wěn)定的轉(zhuǎn)基因表達(dá)。
總結(jié)
通過本文一系列細(xì)胞及使用啟動子的使用情況的整理,我們可以得出如下結(jié)論:
1.CMV啟動子作為最為廣泛使用的啟動子,適合在腫瘤細(xì)胞、肌肉、肝臟等體細(xì)胞或者貼壁細(xì)胞的表達(dá),而不適合大多數(shù)干細(xì)胞、懸浮細(xì)胞及原代細(xì)胞的表達(dá);
2.EF1α啟動子適合干細(xì)胞、原代細(xì)胞、造血細(xì)胞等的表達(dá),在常用細(xì)胞如HEK293、腫瘤等細(xì)胞系中弱與CMV;
3.部分細(xì)胞在傳代過程中,外源啟動子會出現(xiàn)沉默現(xiàn)象,不同的啟動子沉默效率有所差異,對于少見的細(xì)胞系,或者會分化的細(xì)胞系,研究者需要考慮啟動子的選擇,或者多做些分組;
4.CAG啟動子在不少細(xì)胞系,諸如免疫細(xì)胞、體細(xì)胞,有不次于CMV的表達(dá)效率,也是基因表達(dá)的一個較優(yōu)選擇的啟動子。
實(shí)際上,常用于表達(dá)外源基因的啟動子主語局限于CMV、EF1α、CAG、UBC等,本文涉及的PGK、SV40等主要用于表達(dá)熒光、抗性等,很多載體出現(xiàn)熒光弱等問題,可以看看表達(dá)熒光用的何種啟動子。當(dāng)然,諸如懸浮細(xì)胞還可以使用SFFV等啟動子實(shí)現(xiàn)高表達(dá)(可參考吉凱基因懸浮病毒載體使用手冊)。總之,啟動子的選擇對于基因的表達(dá)至關(guān)重要,吉凱也開發(fā)了更加高效表達(dá)的啟動子系列,對于基因表達(dá)弱,熒光弱的老師們,快來吉凱咨詢吧!!!

【參考文獻(xiàn)】
1.Systematic Comparison of Constitutive Promoters and the Doxycycline-Inducible Promoter
2.Quantitative Comparison of Constitutive Promoters in Human ES cells
3.Optimization of adenovirus serotype 35 vectors for efficient transduction in human hematopoietic progenitors: comparison of promoter activities
4.Optimal promoter usage for lentiviral vector-mediated transduction of cultured central nervous system cells
5.Lentiviral Vector Design for Optimal T Cell Receptor Gene Expression in the Transduction of Peripheral Blood Lymphocytes and Tumor-Infiltrating Lymphocytes