?前言
距離全球首例豬心臟移植手術(shù)已經(jīng)過去半年,“頂流”馬里蘭大學(xué)醫(yī)學(xué)院Dr. Deqiang Li的研究團隊于2022年7月在Circulation Research雜志上發(fā)表題目為“Epicardial HDAC3 promotes myocardial growth through a novel microRNA pathway”的心血管文章。該研究揭示了心臟發(fā)育中心外膜HDAC3通過抑制miR-322/miR-503來刺激Fgf9和Igf2進而促進致密心肌生長,為闡明先天性心臟缺陷的病因和促進心肌再生提供了新的策略和見解。
背景介紹
先天性心臟病CHD仍是世界范圍內(nèi)最常見的出生缺陷。肌源性缺陷常與多種形式的冠心病有關(guān)。既往的研究集中在確定心肌的內(nèi)在因素來了解潛在的致病原因,而非心肌細(xì)胞的胞間通訊與調(diào)控作用被忽視。心外膜和心外膜衍生細(xì)胞能夠通過旁分泌信號串?dāng)_調(diào)節(jié)相鄰致密心肌組織的發(fā)育,又通過提供旁分泌生長因子(Fgf9、Igf2)在調(diào)節(jié)心臟再生中起著關(guān)鍵的樞紐作用。
HDAC(組蛋白去乙酰化酶)3是一類HDAC家族的成員,它可以催化從組蛋白尾部賴氨酸殘基中去除乙酰基,并通過調(diào)節(jié)基因表達參與許多生物學(xué)過程。中胚層或全基因敲除HDAC3可導(dǎo)致肌源性缺陷和早期胚胎致死,心肌中特異性的HDAC3缺失卻不會在早期心臟發(fā)育中對心臟形態(tài)表型產(chǎn)生影響,而是會損害出生后后期的心臟功能。這表明HDAC3在非肌細(xì)胞室間的功能可能對早期心肌發(fā)育至關(guān)重要。本研究旨在探尋早期心臟發(fā)育過程中,HDAC3在心外膜中的作用以及Fgf9和Igf2的表達如何受到HDACs或miRs的調(diào)控。
?
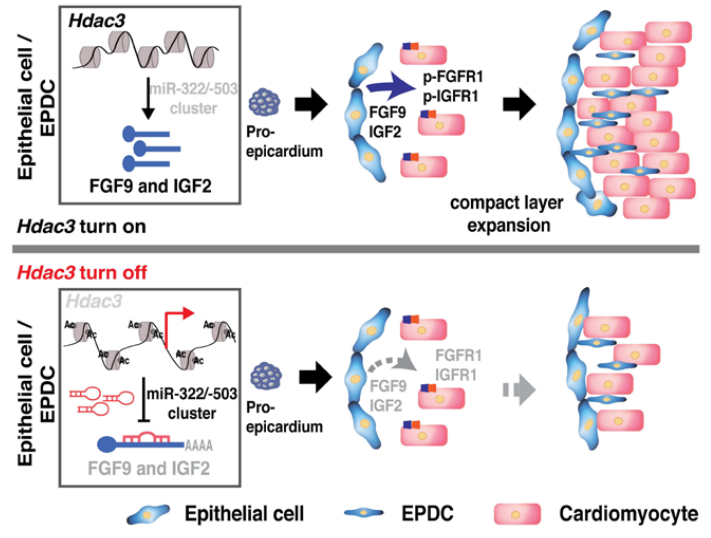
圖. The schematics of the working model
亮點要素
1、發(fā)育中的心外膜中Hdac3的表達對致密心肌生長至關(guān)重要
2、HDAC3對于心外膜衍生的細(xì)胞分化和遷移是很重要的
3、miR-322和miR-503抑制Fgf9和Igf2的轉(zhuǎn)錄
4、HDAC3通過抑制miR-322和miR-503的表達來促進Fgf9和Igf2的轉(zhuǎn)錄
?
研究內(nèi)容
01
研究者首先構(gòu)建了心外膜特異性Hdac3敲除鼠Hdac3eko以研究心臟發(fā)育中Hdac3在心外膜中的潛在作用。對敲除Hdac3的心外膜進行了譜系追蹤。發(fā)現(xiàn)E12.5-E16.5,epdc(心外膜衍生細(xì)胞)發(fā)生EMT遷移至致密心肌成為成纖維細(xì)胞和冠狀動脈血管細(xì)胞。E14.5的eko心臟遷移至致密心肌的epdc比例顯著降低,這表明在Hdac3缺乏的epdc中存在潛在的EMT遷移缺陷。心外膜促進胚胎冠狀動脈發(fā)展,而eko心臟中冠狀動脈血管細(xì)胞標(biāo)志物Sox17陽性細(xì)胞明顯減少,表明eko心臟中冠狀動脈發(fā)育受損。心外膜和epdc通過胞間相互作用促進心肌致密化生長,研究者評估了胚胎期心外膜Hdac3缺陷心臟的心肌,E13.5/E14.5 eko心臟的致密層明顯比對照心臟薄。又對細(xì)胞增殖/凋亡是否導(dǎo)致eko心室壁發(fā)育不良進行驗證,E13.5 eko小鼠致密心肌中pH3/Brdu陽性心肌細(xì)胞的百分比與對照心肌相比顯著降低,證明心臟發(fā)育中心外膜Hdac3特異性缺失導(dǎo)致心室壁發(fā)育不全。
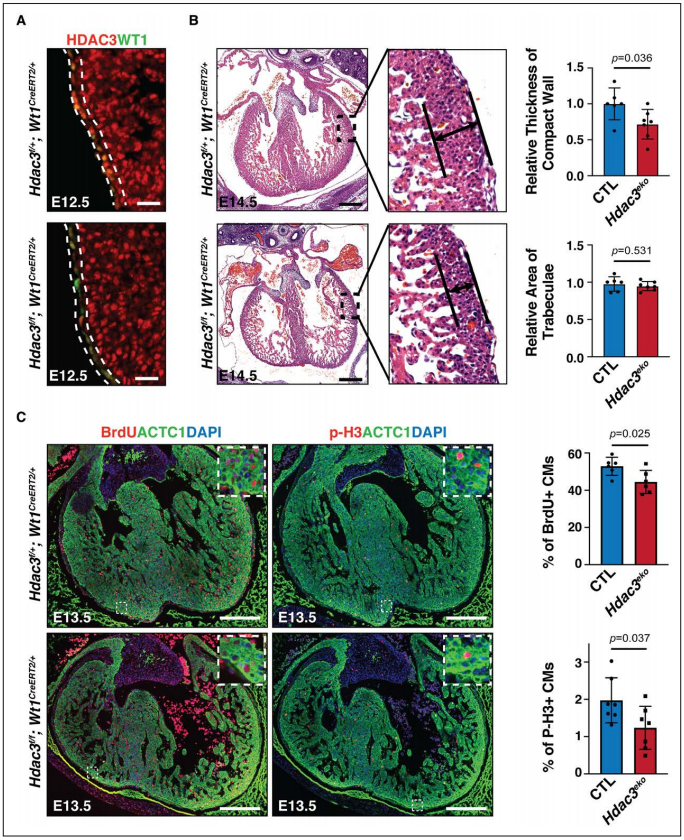
Figure 1. Epicardial deletion of Hdac3 resulted in hypoplasia of ventricular compact wall.
?
?
?
02
為了進一步探究導(dǎo)致eko心臟室壁發(fā)育不全表型的潛在分子機制,研究者使用CRISPR/Cas9技術(shù),構(gòu)建了永生化小鼠心外膜細(xì)胞(MECs)Hdac3單克隆敲除株。Hdac3敲除MECs比對照MECs生長慢。又對細(xì)胞進行批量RNA測序和基因本體分析,發(fā)現(xiàn)了1681個下調(diào)基因。GO通路分析發(fā)現(xiàn),前14個顯著通路均參與細(xì)胞分裂。在分析下調(diào)基因中epdc分泌的生長因子后,確定Fgf9和Igf2是目標(biāo)分子。驗證發(fā)現(xiàn)Hdac3敲除株中Fgf9和Igf2在mRNA和蛋白水平上均降低。體內(nèi)eko心臟中Fgf9和Igf2也顯著降低。提示Fgf9和Igf2的表達下降與Hdac3缺失有關(guān)。
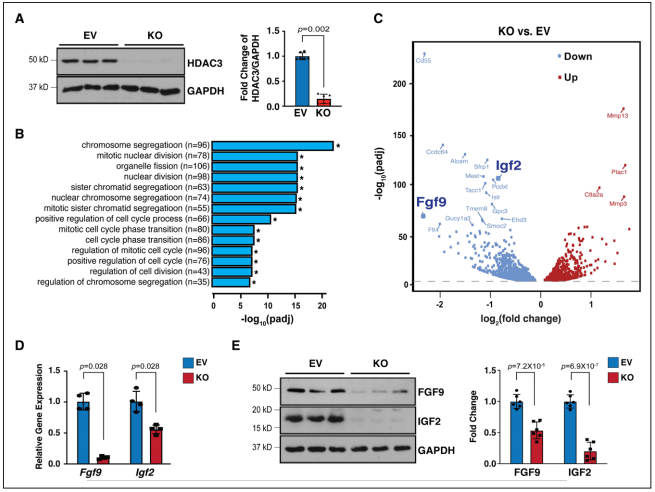
Figure 2. Hdac3 deletion resulted in downregulation of Fgf9 and Igf2.
?
?
03
研究者試圖確定Fgf9和Igf2的減少是否與心肌細(xì)胞增殖減少有關(guān)。與對照細(xì)胞EV相比,Hdac3敲除株KO上清液中Fgf9和Igf2顯著降低。又用細(xì)胞上清處理E13.5分離的原代胚胎心肌細(xì)胞。與EV上清相比,KO上清處理的pH3陽性心肌細(xì)胞百分比和心肌細(xì)胞總數(shù)顯著降低并且其減弱了心肌細(xì)胞增殖下游信號通路ERK磷酸化的激活。而在KO中補充Fgf9和Igf2可以修復(fù)心肌細(xì)胞增殖缺陷并使p-FGFR1/p-IGFR1活化。以上結(jié)果表明,來自心外膜細(xì)胞分泌的Fgf9和Igf2提供了驅(qū)動心肌細(xì)胞增殖的重要線索。
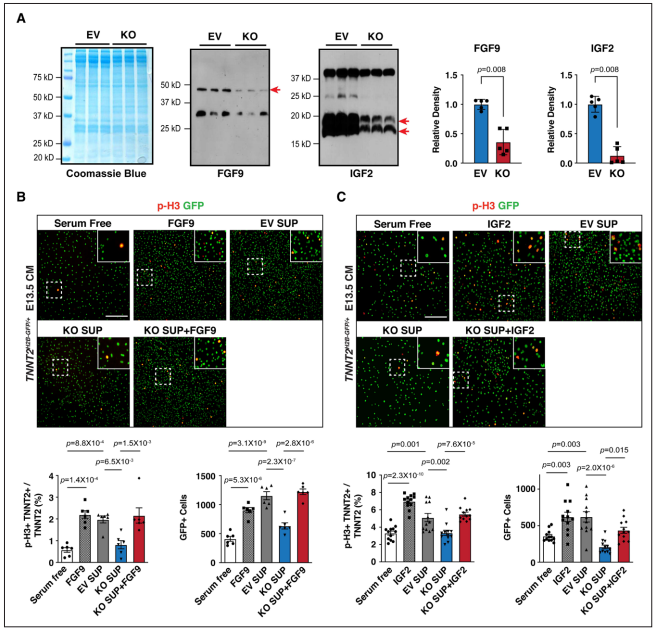
Figure 3. Supplementation of Fgf9 or Igf2 rescues CM proliferation defects.
?
?
?
04
為了確定HDAC3是通過去乙酰化酶活性來誘導(dǎo)Fgf9和Igf2的表達,研究者用選擇性HDAC3去乙酰化酶抑制劑RGFP966處理MECs,RGFP966處理顯著降低了Fgf9和Igf2。研究者又進行了Rescue實驗。慢病毒Hdac3在敲除株中重表達成功恢復(fù)了Fgf9和Igf2的表達,而去乙酰化酶失活突變慢病毒Hdac3的重表達并不能挽救Fgf9和Igf2的表達。證明HDAC3確實是通過去乙酰化酶活性調(diào)控Fgf9和Igf2的表達。
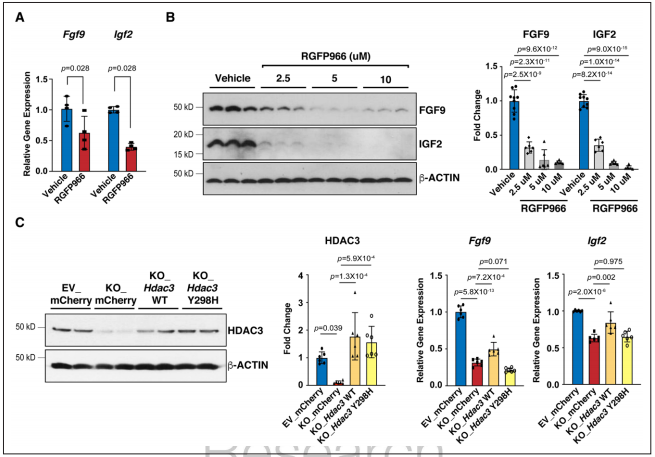
Figure 4. HDAC3 induces the expression of Fgf9 and Igf2 dependent on its deacetylase activity.
?
?
?
05
Fgfs和Igfs的表達可被miRs調(diào)控,研究者對敲除株進行了miR測序。通過差異表達分析及模擬結(jié)合位點篩選發(fā)現(xiàn)miR-322或miR-503模擬物處理顯著抑制Fgf9和Igf2的表達。通過Western blot進一步驗證了miR-322 mimics/miR-503 mimics處理對Fgf9和Igf2表達的顯著抑制作用。又使用miR-322或miR-503模擬物處理培養(yǎng)的E13.5心肌細(xì)胞,顯著降低了pH3陽性心肌細(xì)胞的百分比,表明miR-322和miR-503抑制心肌細(xì)胞增殖。當(dāng)HDAC3的去乙酰化酶活性被RGFP966抑制時,miR-322和miR-503與敲除株中同樣顯著上調(diào)。體內(nèi)E13.5 eko心臟中也觀察到miR-322和miR-503的顯著上調(diào)。證明了miR-322、miR-503能夠抑制Fgf9、Igf2表達和心肌細(xì)胞增殖。
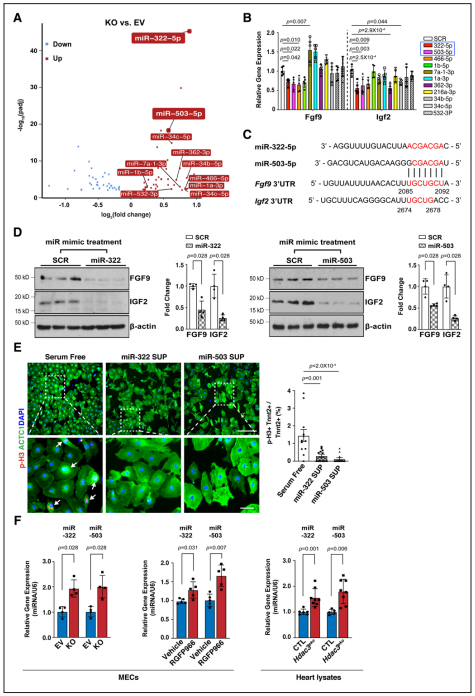
Figure 5.miR-322 and miR-503 repress the expression of Fgf9 and Igf2 and CM proliferation.
?
06
為了確定當(dāng)Hdac3被敲除或抑制時,miR-322和miR-503的上調(diào)對Fgf9和Igf2的降低有因果影響,使用miRZip慢病毒干擾miR-322或miR-503的表達,敲低miR-322或miR-503后可顯著恢復(fù)敲除株中Fgf9和Igf2的表達。以上結(jié)果表明HDAC3通過抑制miR-322和miR-503促進Fgf9和Igf2的表達。

Figure 6. Knockdown of miR-322 or miR-503 restores the expression of Fgf9 and Igf2.
?
07
H3K27Ac是活性啟動子的標(biāo)記,也是HDAC3的直接下游靶點。Hdac3缺失導(dǎo)致MECs中H3K27Ac顯著增加。ENCODE實驗發(fā)現(xiàn),miR-322/miR-503的啟動子區(qū)域受表觀遺傳調(diào)控。為了確定Hdac3缺失是否會影響miR-322/miR-503啟動子的染色質(zhì)可及性,研究者通過ChIP-qPCR研究了H3K27Ac在敲除株中的結(jié)合親和力。發(fā)現(xiàn)Hdac3缺失顯著增加了H3K27Ac與miR-322/miR-503啟動子的結(jié)合,這表明Hdac3缺失使miR-322/miR-503啟動子更容易被轉(zhuǎn)錄因子接近。為了明確這種染色質(zhì)可及性的增加是否會影響miR-322/miR-503啟動子的活性,又進行了miR-322/miR-503啟動子熒光素酶報告基因?qū)嶒灐0l(fā)現(xiàn)與對照細(xì)胞株相比,敲除株或經(jīng)RGFP966處理后熒光素酶活性顯著增加,表明HDAC3對miR-322/miR-503啟動子的調(diào)控同樣依賴于其去乙酰化酶活性。
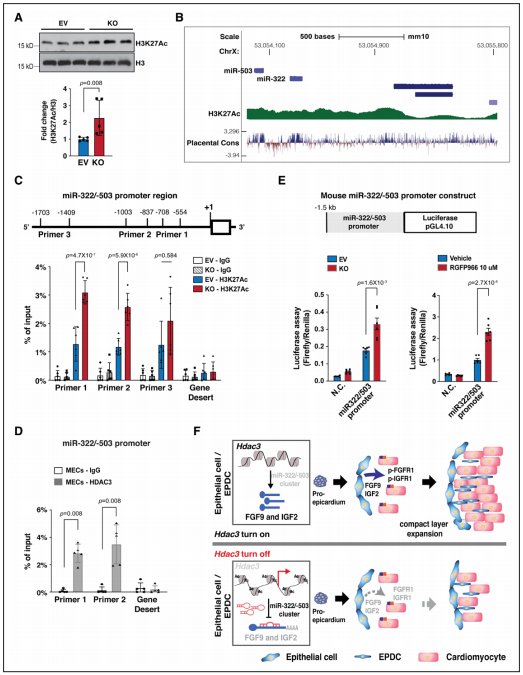
Figure 7. HDAC3 represses miR-322/miR-503 promoter activity.
?
解暑好物!心血管及代謝疾病動物模型整體實驗8折優(yōu)惠!夏日解憂實驗室!
?
和元生物具有多年豐富的動物實驗和心血管動物模型構(gòu)建的實操經(jīng)驗,并兼具出色的基因操作與病毒構(gòu)建等技術(shù)。同時依托公司的檢測平臺、病理平臺、細(xì)胞平臺等專業(yè)實驗平臺,可為您提供從心血管動物模型到基因干預(yù)、病理檢測、細(xì)胞功能學(xué)研究等一站式心血管研究服務(wù)體系,您可體驗文中涉及到的心血管前沿研究手段與支持。歡迎關(guān)注和元公眾號了解實驗服務(wù)詳情,和元生物期待您的到來。