
期刊:Nature Communications
影響因子:14.7
主要技術(shù):scRNA-seq、Xenium
導語
研究人類胎兒肺可以了解發(fā)育缺陷和疾病狀態(tài)如何改變肺的功能。在這里,我們對19個健康人假腺體胎兒肺組織的>150000個單細胞進行了測序,時間范圍在妊娠10-19周之間。我們從表達豐富水平的囊性纖維化電導跨膜調(diào)節(jié)因子 (CFTR)的祖細胞中捕獲動態(tài)發(fā)育軌跡。這些細胞產(chǎn)生多種特化的上皮細胞類型。結(jié)合空間轉(zhuǎn)錄組學,我們展示了關(guān)鍵信號通路的時間調(diào)控,這些通路可能驅(qū)動特化上皮細胞(包括纖毛細胞和肺神經(jīng)內(nèi)分泌細胞)的時間和空間出現(xiàn)。最后,我們表明人類多能干細胞衍生的胎肺模型包含表達 CFTR 的祖細胞,這些祖細胞捕獲與天然組織中鑒定的相似的譜系發(fā)育軌跡。總體而言,本研究提供了發(fā)育中人肺的全面單細胞圖譜,概述了細胞譜系發(fā)育的時間和空間復雜性,并對從人多能干細胞分化到類似發(fā)育窗口的胎兒肺培養(yǎng)進行了基準測試。
主要技術(shù)
scRNA-seq、Xenium
研究結(jié)果
1.人胚胎肺的細胞組成
收集總共19個來自選擇性終止妊娠的新鮮分離的胎兒肺組織,范圍從孕周(GW)10-19,并進行處理,用于3 '單細胞RNA測序(scRNA-seq)。從肺部的不同區(qū)域?qū)σ粋€組織進行兩次采樣,以確保均勻捕獲。總共對170256個單細胞進行了測序,每個細胞的讀取深度約為60000。生成FASTQ文件,并將讀數(shù)與人類參考基因組hg 38(GRCh 38)進行比對。去除低質(zhì)量細胞、死細胞和雙細胞后,分析了156,698個細胞。經(jīng)過標準化和降維后,使用倒數(shù)主成分分析(rPCA)來識別錨點,以進行整合以生成胎兒肺數(shù)據(jù)集。根據(jù)男性性別決定基因SRY和DDX 3 Y以及XIST的表達,收集了12個男性和7個女性的肺部(圖1A)。使用基于非參數(shù)Wilcoxon等級和檢驗的差異表達基因(DEG)來注釋細胞群,將細胞群廣泛分為5個獨特不同的細胞群(圖1B)指定為矩陣(N = 98166; Col 1A 1、Col 1A 2),表皮(N = 16068; EPCAM)、內(nèi)皮細胞(N = 13376; PECAM 1)、免疫細胞(N = 16258; CD 74、CD 3、NKG 7)和施旺細胞(N = 512; NRXN 1、S100 B、PLP 1)(圖1C)。Clustree和DEG對每個主要細胞群進行了進一步的亞聚集,這導致發(fā)育中的肺部中總共有58種不同的細胞類型/狀態(tài)(圖1D)。評估整個妊娠時間的細胞比例顯示,在整個10周的發(fā)育過程中,間質(zhì)細胞構(gòu)成了最大的細胞比例(圖1 E)。
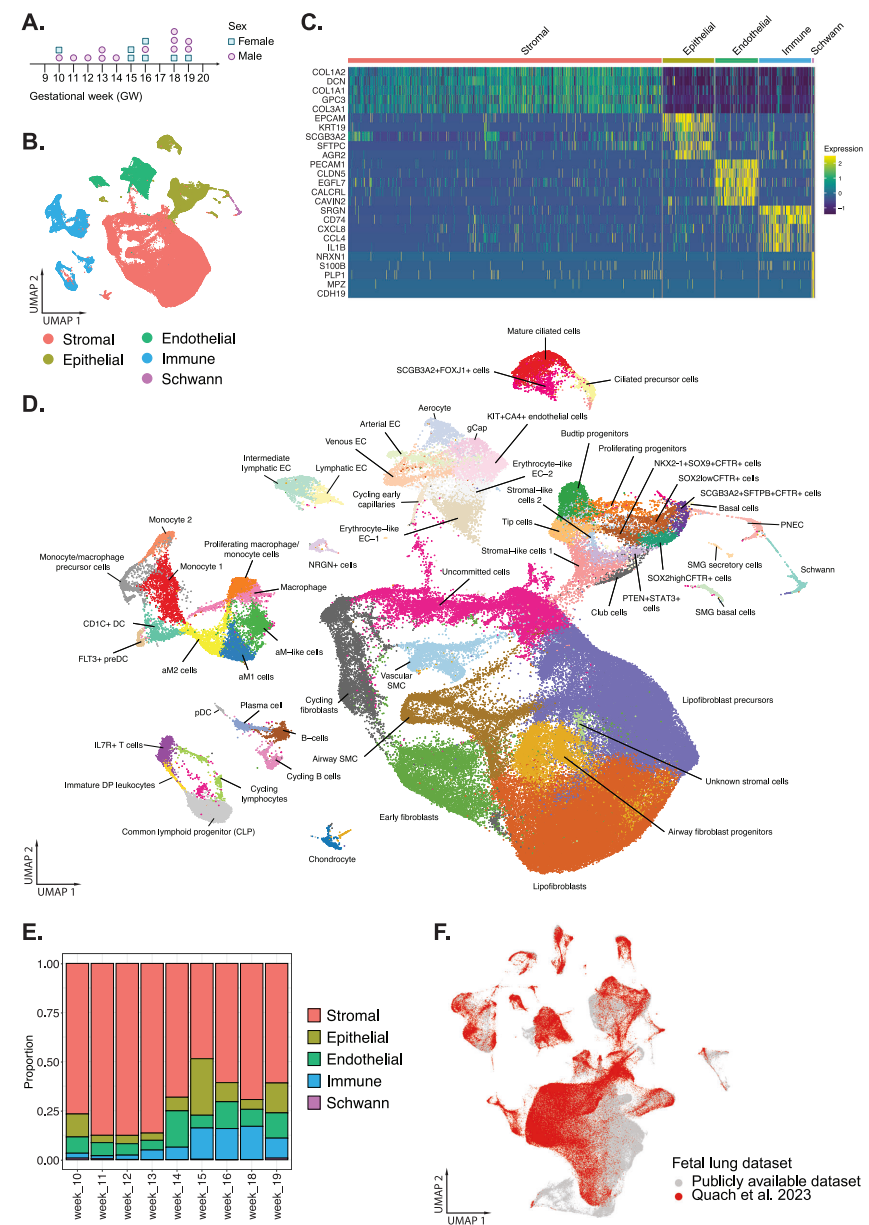
圖1
2. 發(fā)育中的肺間質(zhì)
在分析的所有肺組織中,間質(zhì)細胞構(gòu)成了最大比例的細胞(圖1B)。我們根據(jù)DEG確定了10種間質(zhì)細胞亞型(圖2A)。這些包括脂成纖維細胞、脂成纖維細胞前體、循環(huán)成纖維細胞、早期成纖維細胞、氣道成纖維細胞祖細胞、未定型細胞、氣道平滑肌細胞、血管平滑肌細胞、軟骨細胞和未知的間質(zhì)細胞群。脂成纖維細胞和脂成纖維細胞前體是整個妊娠周中含量最豐富的間質(zhì)細胞類型(圖2B)。這兩種細胞類型都表達TCF 21,這是一種先前在小鼠胎兒和成人肺成脂纖維細胞中顯示的標志物。脂肪成纖維細胞前體差異表達更高水平的幾種典型間充質(zhì)相關(guān)基因,包括PLEKHH 2和MACF 1(圖2C),類似于小鼠脂肪成纖維細胞細胞系。在脂肪成纖維細胞中鑒定的最高差異轉(zhuǎn)錄因子JUN、FOS和STAT 3的調(diào)節(jié)子活性(圖2D),已知其調(diào)節(jié)結(jié)構(gòu)性細胞外基質(zhì)(如膠原蛋白和彈性蛋白)的合成,并且在肺泡發(fā)育中很重要。雖然肺泡發(fā)育發(fā)生在妊娠后期,但早期胎兒肺發(fā)育中的脂成纖維細胞在形成氣道結(jié)構(gòu)所需的ECM蛋白中發(fā)揮作用。驅(qū)動脂肪成纖維細胞前體的轉(zhuǎn)錄因子的差異調(diào)節(jié)子活性包括NR2F1和ETV 1,其調(diào)節(jié)細胞生長、繁殖和分化(圖2D)。氣道成纖維細胞表達高水平的TWIST 1、NIX和ZEB 1(圖2D)。
接下來,我們使用Xenium高分辨率多路原位空間分析來確定每個間質(zhì)細胞亞型中表達的DEG的空間定位。我們在胎兒肺組織的間質(zhì)區(qū)域中鑒定了與氣道成纖維細胞祖細胞(WRPINF 1、FBN 1、IGMBE 5)、脂成纖維細胞前體(MACF 1和TCF 21)和脂成纖維細胞(ERG 1、ATF 3、IRF 1)相關(guān)的基因(圖2 E)。氣道成纖維細胞祖細胞主要集中在大氣道(表皮1)周圍,而脂成纖維細胞前體則出現(xiàn)在整個間質(zhì)中。此外,還觀察到分散在表皮周圍的脂成纖維細胞前體之間的脂成纖維細胞。
周期性成纖維細胞和早期成纖維細胞在早期肺組織中也以相對較高的比例存在(GW 10-14;分別為~ 6-10%和~ 9-12%),但在晚期妊娠肺組織中逐漸減少(GW 15;分別為~2-5%和~3-7%)(圖2B)。除了常見的膠原基因(Col 1A 1、Col 1A 2)外,周期成纖維細胞表達了高水平的與細胞增生相關(guān)的基因TOP 2A、CENPF、MKI 67(圖2C)。早期成纖維細胞表達了高水平的與染色質(zhì)重塑相關(guān)的基因,例如HELS和TYMS 29(圖2D),此前已證明這些基因在小鼠胚胎成纖維細胞中高度表達。早期成纖維細胞中具有差異性高調(diào)節(jié)子活性的轉(zhuǎn)錄因子包括E2 F7、E2 F8、FOXM 1、TCF 7和E2 F1。
根據(jù)UMAP預測,還有很大一部分未定型細胞與周期成纖維細胞和脂肪成纖維細胞前體密切相關(guān)(圖2A)。這些未定向的細胞與循環(huán)成纖維細胞和脂肪成纖維細胞前體有一些轉(zhuǎn)錄相似性,但也表達與其他細胞類型相關(guān)的基因(圖2C)。氣道SMC差異表達更高水平的TAGLN、DES和MYH 11,而血管SMC表達IGFBP7、MEF 2C和EGFL 6,這兩種情況在其他研究中也有類似的報道。TBX 5、MYEF 2和KLF 9的轉(zhuǎn)錄因子和相關(guān)調(diào)節(jié)因子在氣道SMC中具有差異活性,而ZFN 41、PITX 3和HMGA 1在血管SMC中存在。軟骨細胞差異表達許多膠原蛋白基因,包括Col 2A1、Col 11 A1、Col 9A1和RUNX 1,以及已知調(diào)節(jié)軟骨細胞發(fā)育的FOXO 1相關(guān)調(diào)節(jié)子。具有明顯DEG的未知一小群間質(zhì)細胞(第10群)表達了高水平的AHSP、ALAS和BPGN,這些基因與紅細胞相關(guān)。
為了推斷成纖維細胞團的發(fā)育譜系,我們使用LatentVelo來估計不含氣道SMC、血管SMC和軟骨細胞的一組間質(zhì)細胞上的RNA速度。為這個子集生成了新的UMAP,我們在UMAP嵌入和PAGA上可視化了這些速度(圖3A、B)。推斷軌跡表明,周期性成纖維細胞形成早期成纖維細胞,然后形成氣道成纖維細胞祖細胞和脂成纖維細胞。我們使用Slingshot進一步驗證了這些軌跡,使用循環(huán)成纖維細胞作為根細胞或細胞起源,并發(fā)現(xiàn)隨著細胞沿著分化軌跡移動為脂成纖維細胞或氣道成纖維細胞祖細胞,基因表達發(fā)生了逐漸變化(圖3C、D)。
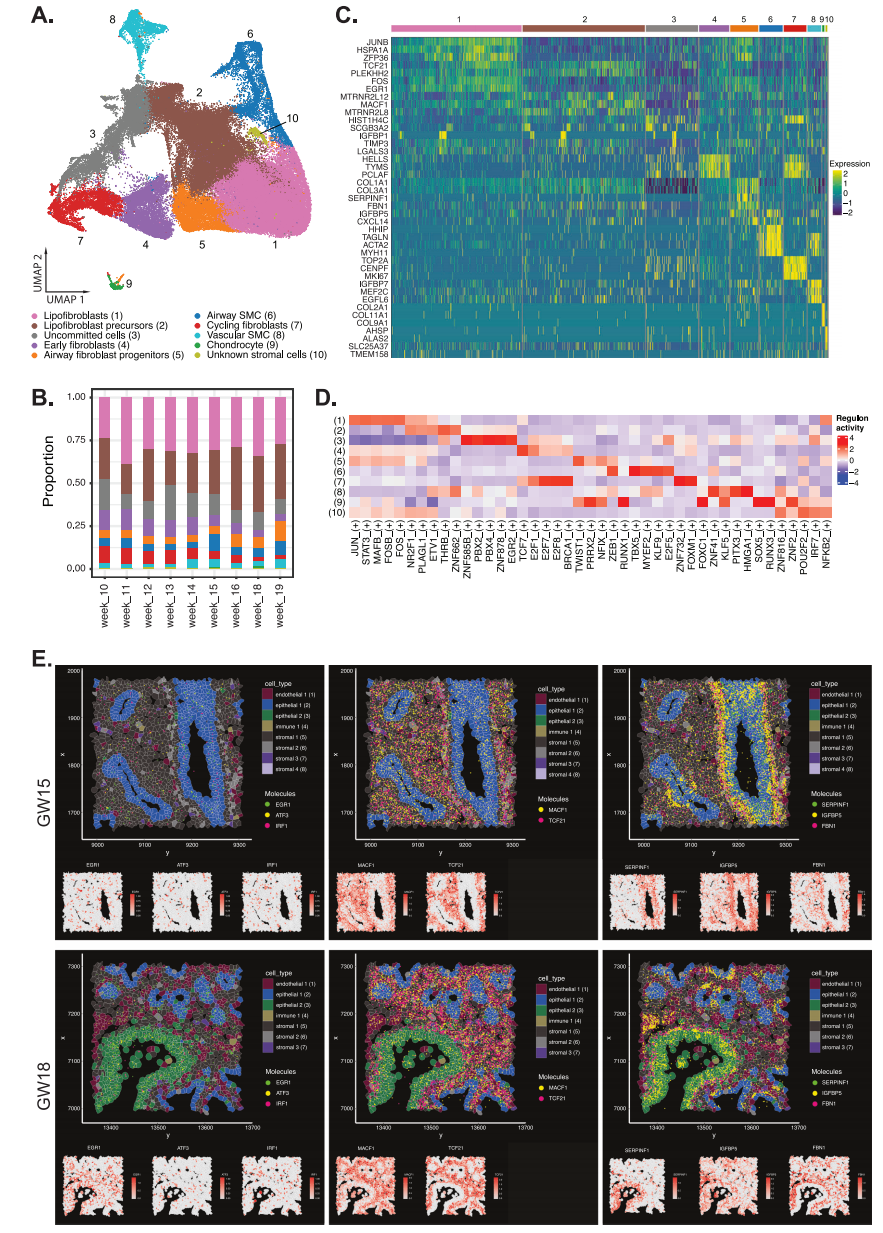
圖2

圖3
3. 動態(tài)演變的胎兒肺細胞
在胎兒肺表皮中,對topDEG的分析加上與特定細胞相關(guān)的典型表皮基因的高表達,揭示了19種亞型(圖4A、B)。利用SOX 2和SOX 9的表達分別確定近端和遠端氣道細胞,我們鑒定了表達高水平SOX 2的表皮細胞亞群,這是一種共表達SCGB 3A2和SFTPB的CFTR表達細胞類型(SCGB 3A2 + SFTPB + CFTR +)、SCGB 3A 2 + FOXJ 1+和SOX 2 highCFTR+祖細胞群(圖4C)。通過與細胞增生相關(guān)的基因MKI 67和編碼微小管結(jié)合蛋白CENPF、NUSAP 1和CDK 1的細胞周期基因的表達升高來鑒定新生祖細胞(圖4 B)。 空間Xenium分析顯示NKX 2 -1 + SOX 9 + CFTR +僅限于表皮(表皮2,藍色陰影),所有三個基因在GW 15和18肺組織的分叉區(qū)域或發(fā)育芽區(qū)均豐富表達(圖4D)。相比之下,在氣道的莖部區(qū)域發(fā)現(xiàn)了MEK 2 + CFTR +細胞。已知SOX 2表達可以調(diào)節(jié)表皮細胞的細胞增生,因此,SOX 2 highCFTR+細胞可能參與細胞擴張以形成發(fā)育中的氣道。
使用免疫熒光染色,我們證實了NKX 2 -1、SOX 9和CFTR三陽性細胞在發(fā)育中的肺芽區(qū)域中的空間表達(圖4 E)。相反,在胎兒氣道中發(fā)現(xiàn)NKX 2 -1 + SOX 2 + CFTR +細胞,具有明顯的SOX 2高(橙色箭頭)和SOX 2低(白色箭頭)細胞群。SCGB 3A 2 + SFTPB + CFTR +細胞在本文中被稱為三重陽性(TP),差異表達更高水平的CXCL17、CP和CYB 5A(圖4 B)以及轉(zhuǎn)錄因子HER 1(主要的NOTCH靶基因)和ASCL 2的差異高調(diào)節(jié)子活性,ASCL 2之前被證明是腸道表皮中的WNT/CTNNB 1轉(zhuǎn)錄靶點。發(fā)現(xiàn)這些TP細胞大量散布在整個氣道中(圖4D、E)。
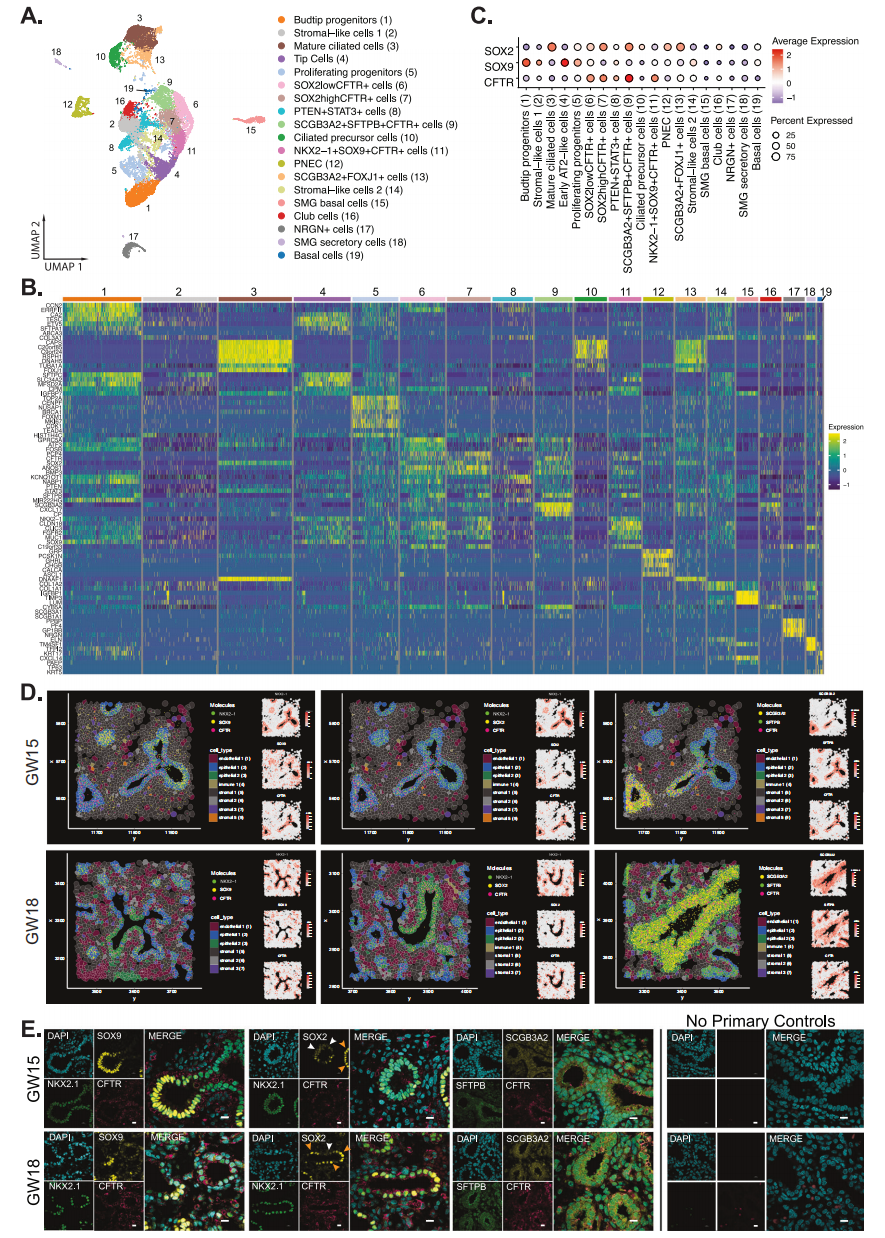
圖4
4. 推測的表皮亞型軌跡
使用LatentVelo估計了RNA速率,以預測表皮細胞類型的譜系關(guān)系(圖5A)。然后,我們使用CellRank 分析了LatentVelo速度,并確定了三種終末狀態(tài):成熟的纖毛細胞、PNEC和芽尖祖細胞(圖5B)。基于使用LatentVelo的PAGA,我們觀察到表明細胞高度可塑性和細胞譜系關(guān)系變化的軌跡(圖5C)。假設(shè)表皮譜系的發(fā)育可能受到時間調(diào)節(jié)。基于幾種典型細胞類型的出現(xiàn),例如成熟的纖毛細胞和GW 14時PNEC細胞的顯著下降(圖5D)。在早期妊娠組織中,PNEC的貢獻有多種來源,包括從芽尖祖細胞到尖端細胞到SOX 2 highCFTR到TP,然后是PNEC的軌跡。DEG的變化反映了細胞命運逐漸改變?yōu)镻NEC分化(ASCL 1和GHRL的表達)(圖5F)。沿著軌跡的細胞的富集反映了表型的變化。在從TP細胞向PNEC轉(zhuǎn)化的過程中,參與FGF、NOTCH和TGFβ信號通路的基因上調(diào)。所有的PNEC軌跡在后期減弱,并且在這些后期階段的PNEC細胞的比例降低(圖5D)。幾條軌跡聚集在妊娠中期組織中的TP上(圖5E)。這些包括來自SOX 2lowCFTR+細胞、SOX 2 highCFTR+細胞、間質(zhì)樣細胞、增生祖細胞和基礎(chǔ)細胞的細胞來源。在所有時間點都觀察到NKX 2 -1 + SOX 9 + CFTR +和SOX 2lowCFTR+細胞之間的強聯(lián)系(粗箭頭線),以及后兩種細胞具有TP,表明這些細胞與發(fā)育相關(guān)。我們推斷的軌跡預測(圖5F、G)和Xenium空間定位表明了SOX 9+細胞之間的關(guān)系。

圖5
5. PNEC和纖毛細胞譜系中TP細胞的可塑性
離體譜系追蹤研究表明,TP細胞可以在功能上對PNEC和下呼吸道中的纖毛細胞做出貢獻。在我們的數(shù)據(jù)集中,我們發(fā)現(xiàn)這些細胞后期組織中PNEC下降,GW 14周圍的纖毛細胞隨之增加(圖6A)。CellRank用于估計TP細胞的PNEC和成熟纖毛細胞命運概率,顯示隨著GW的增加,PNEC概率下降,成熟纖毛細胞概率增加(圖6B)。PNEC標志物ASCL 1(黃色)和纖毛蟲細胞標志物FOXJ 1(品紅色)的免疫熒光染色顯示早期GW組織中富含PNEC細胞,而這些細胞在GW 18胎肺中丟失(圖6C)。空間轉(zhuǎn)錄組學的共現(xiàn)分析顯示,這兩種細胞類型都靠近TP細胞和SOX 2+細胞,表明這些細胞位于發(fā)育中的莖區(qū),并對氣道的譜系組成做出貢獻(圖4A、B)。總而言之,這些結(jié)果證明了PNEC和通過TP細胞的纖毛細胞分化的時間調(diào)節(jié),支持這些祖細胞在發(fā)育中的肺中的動態(tài)可塑性。
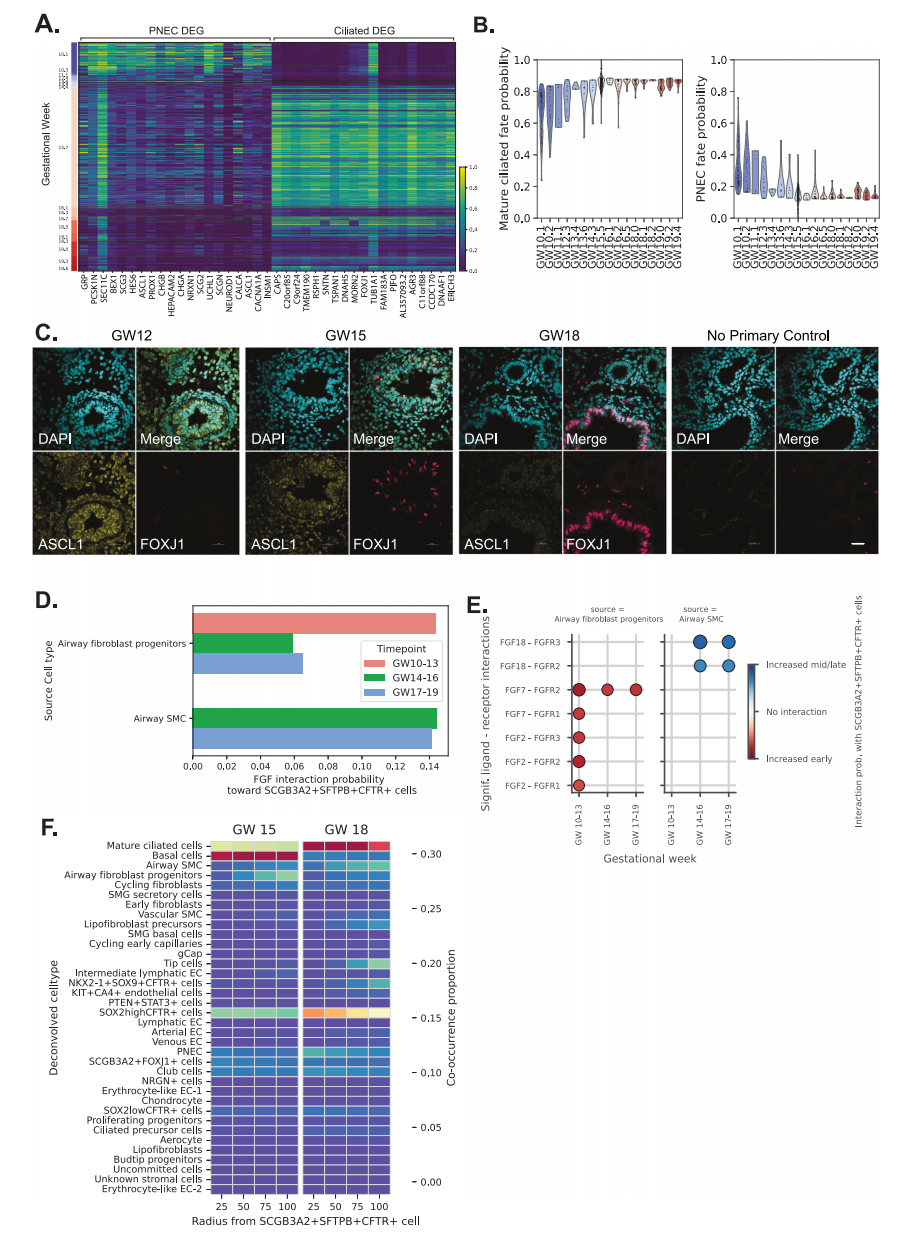
圖6
6. FGF信號傳遞至TP的時間變化
相對于其他表皮細胞特異性靶向TP細胞的信號途徑包括成纖維細胞生長因子(FGF,圖6D)。FGF信號途徑是一種參與肺分支形態(tài)發(fā)生的已知途徑。此外,在定向分化方案中使用了幾種FGF重組蛋白,以從人iPSC 16、18產(chǎn)生胎兒肺表皮細胞。因此,我們推測該途徑可能在調(diào)節(jié)TP細胞命運方面發(fā)揮著重要作用。我們特別關(guān)注通過FGF信號與TP細胞的細胞間相互作用(圖6 D)。根據(jù)Xenium的信息,我們鑒定了在空間上與TP細胞“緊密接近”的細胞,其中包括GW 15肺中的氣道成纖維細胞祖細胞、SOX 2 highCFTR+細胞、PNEC、Club細胞和基礎(chǔ)細胞,以及脂成纖維細胞前體、SCGB 3A 2 + FOXJ 1+、氣道SMC和GW 18肺中的成熟纖毛細胞(圖6 E)。然后,我們估計了參與TP細胞和FGF信號之間的特定配體-受體(L-R)相互作用。我們專門研究了氣道成纖維細胞祖細胞和氣道SMC,因為它們與基于共存細胞類型的TP細胞非常接近(圖6E)。我們發(fā)現(xiàn),妊娠早期(GW 10 -13)期間,涉及氣道成纖維細胞祖細胞中表達的配體FGF 2和FGF 7的L-R相互作用之間的信號傳遞顯著增加,并且我們發(fā)現(xiàn),涉及氣道SMC中表達的配體FGF 18的L-R相互作用的信號傳遞顯著增加(GW 14 -16)或晚期(GW 17 -19)(圖6F)。
7. hPSC衍生的胎兒肺分化捕獲細胞異質(zhì)性和在原生組織中發(fā)現(xiàn)的軌跡
hPSC的定向分化方案旨在捕捉發(fā)育里程碑,確保在細胞培養(yǎng)中真實細胞類型的穩(wěn)健發(fā)展。隨著分化變得更加先進,有能力產(chǎn)生hPSC衍生的組織特異性類器官,這些模型代表了研究發(fā)展基本機制。當獲得用于研究的主要胎兒組織受到限制時,這一點尤其重要。在這里,我們試圖使用我們最新的肺分化方案來確定我們認為是“胎兒”肺細胞和由hPSC產(chǎn)生的類器官的細胞的發(fā)育階段(圖7A)。我們對hPSC衍生的胎兒肺細胞進行了sc-RNA測序,認為其包含大部分未分化的肺上皮細胞和胎兒類器官,這些細胞將沿著分化途徑進行下一步。
在聚類并進行DEG分析后,我們將這些hPSC細胞簇的DEG與原代胎兒肺樣本中的DEG進行了比較。在hPSC胎兒肺細胞中,我們發(fā)現(xiàn)簇0和1中的增殖祖細胞和芽尖祖細胞有明顯的重疊(圖7B,C)。在這些胎兒肺細胞培養(yǎng)中出現(xiàn)的其他細胞類型的DEG包括PNEC、SOX2+、SOX9+祖細胞和第2簇的TP細胞。相反,hPSC衍生的胎兒肺類器官,代表了下一階段的分化,包含與基底細胞、club細胞、PNEC、SMG分泌細胞、纖毛細胞和TP細胞顯著重疊的細胞(圖7D,E)。集群0和集群2中基底細胞的高度重疊并不令人驚訝,因為我們的類器官擴增培養(yǎng)基含有雙重TGFβ/SMAD信號抑制劑:DMH1和A83-01,它們旨在促進基底細胞擴增。
為了將這些hPSC分化細胞與胎兒肺發(fā)育的妊娠時間點進行比較,我們使用了Spearman相關(guān)系數(shù),發(fā)現(xiàn)hPSC分化細胞與這些早期胎兒肺組織的相關(guān)性相對較強。我們發(fā)現(xiàn)hPSC衍生的胎兒肺細胞與<GW12肺上皮組織相關(guān)性最強,hPSC衍生的胎兒肺類器官與>GW16肺上皮組織相關(guān)性最強(圖7F)。重要的是,沒有一種hPSC胎兒肺模型與成人肺有很強的相關(guān)性。
使用鄰域分析,我們整合了hPSC來源的細胞和原代胎兒肺上皮(圖8A)。綜合UMAP顯示,hPSC衍生的胎兒肺細胞(紫色)和類器官(綠色)與原代胎兒肺組織中的上皮細胞有明顯的重疊(圖8B)。為了了解這些hPSC分化在多大程度上代表了胎兒肺上皮的發(fā)育,我們將這些細胞定位在我們之前從天然組織推斷的軌跡上。我們考慮了先前推斷的胚胎初代上皮從芽尖祖細胞到TP細胞,通過尖端細胞,NKX2-1 + SOX9 + CFTR +細胞和sox2高/低CFTR +細胞的軌跡(圖8C)。我們使用集成的最近鄰圖對集成的細胞進行聚類,并且只評估沿軌跡至少有100個細胞的聚類。因此,分析中沒有顯示胎兒和hPSC來源的培養(yǎng)物之間細胞重疊<100個細胞的集群(圖8D)。
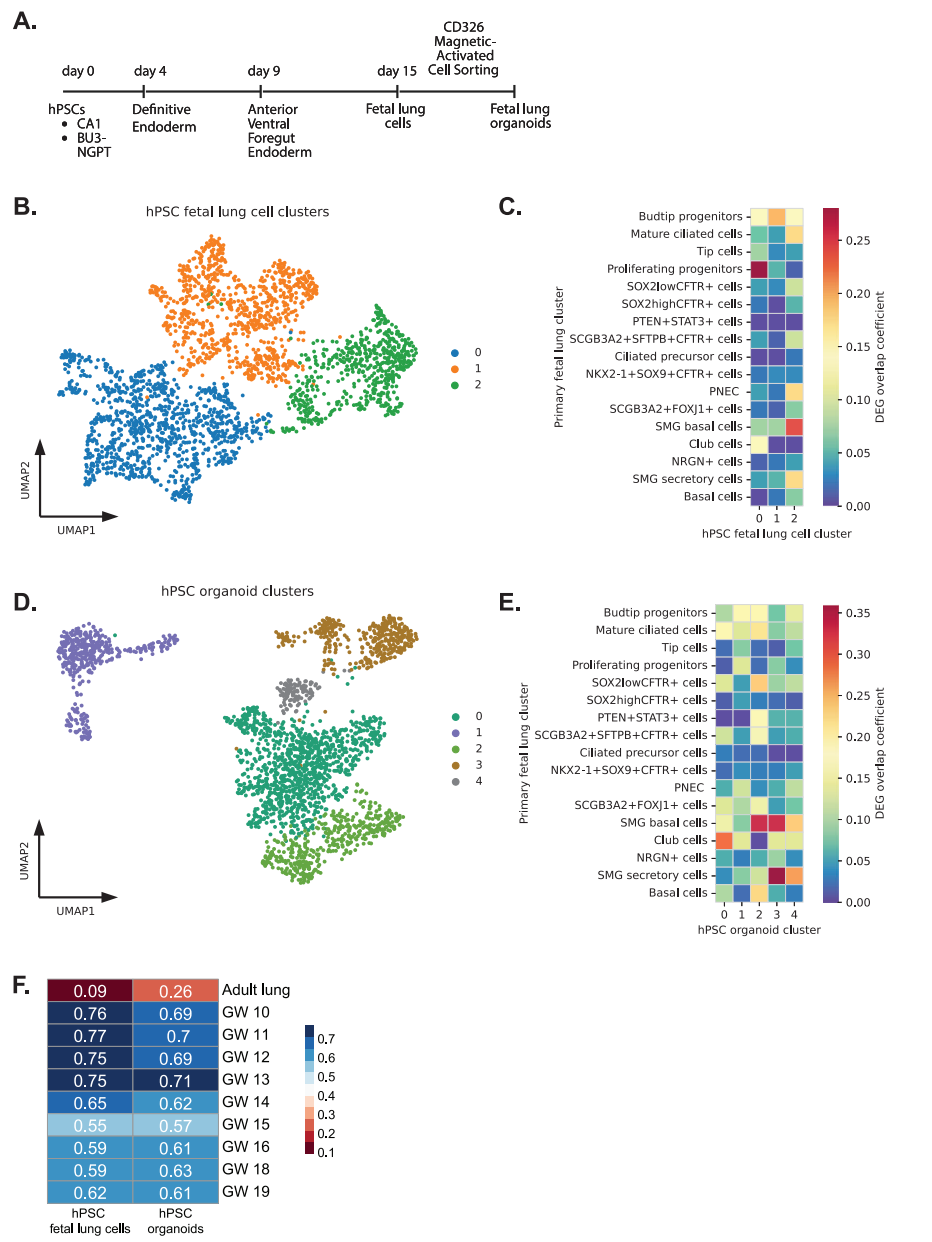
圖7

圖8
為了不偏不倚地確定hPSC衍生培養(yǎng)物中沿軌跡的上皮細胞亞型,我們從原代上皮組織中鑒定的每個簇的top100DEG中創(chuàng)建細胞類型評分,并使用每個包含hPSC胎兒肺細胞和類器官細胞的這些簇的平均擬時序,我們沿著軌跡定位簇(圖8E,上面的箭頭指向軌跡方向)。這些分數(shù)表明,隨著獲得與各種分化上皮細胞類型/狀態(tài)(如基底細胞和TP細胞)相關(guān)的特定基因,胎兒肺類器官沿著這條軌跡逐漸分化。在hPSC胎兒肺細胞簇中,增殖祖細胞類型評分隨著擬時序的增加而增加,而早期hPSC類器官簇隨著增加而消失,而類器官中sox2高/低cftr +、TP和基底細胞類型評分則增加。基因表達變化的清晰模式沿著平均擬時序出現(xiàn),在hPSC胎兒肺細胞中可以看到增殖祖標記基因的缺失(如TOP2A和CENPF),而獲得了鑒定分化程度更高的細胞類型的基因,如sox2高/低cftr +細胞的GPC5A, TP細胞的CP和基底細胞的SPRR3(圖8F)。
為了進一步證實整合細胞的分析,我們使用LatentVelo對hPSC胎兒肺細胞和hPSC類器官進行了獨立的RNA速率分析。與綜合軌跡分析類似,我們發(fā)現(xiàn)從hPSC胎兒肺集群0到集群1和2的軌跡(圖8G),以及從hPSC類器官集群1和4到集群0和2的軌跡(圖8H)。
總體而言,我們的hPSC分化胎兒細胞模型捕獲了一些上皮細胞類型/狀態(tài)和軌跡,特別是從芽尖到TP細胞,以及在原代胎兒假腺/小管肺組織中觀察到的幾種中間狀態(tài)(圖8I)。
總之,我們提供了人類胎兒肺的高分辨率時空圖譜,重點關(guān)注上皮細胞和相互作用的基質(zhì)細胞在形成發(fā)育中的氣道中的譜系關(guān)系(圖9)。
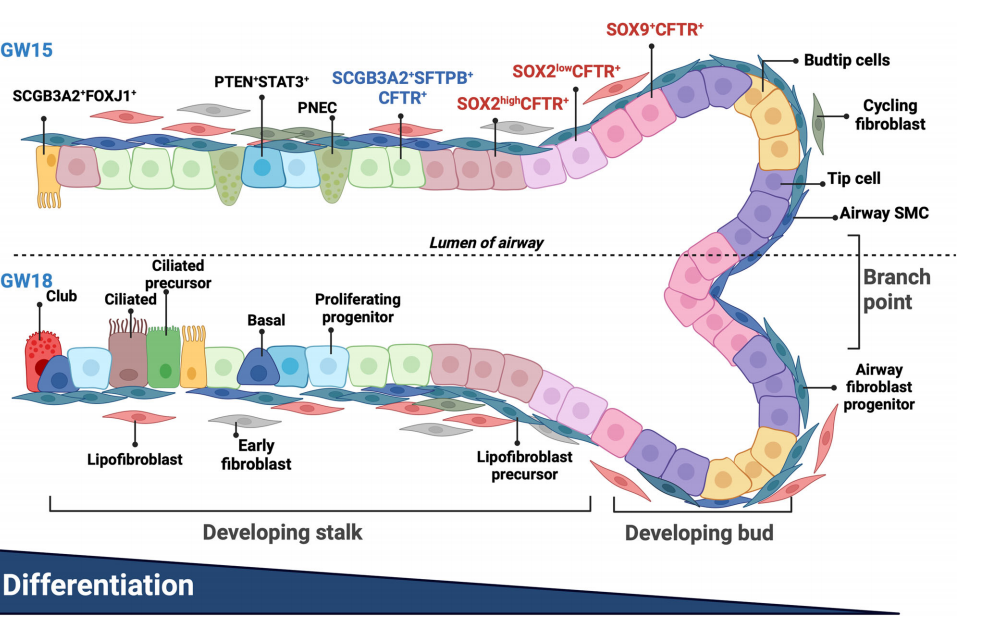
圖9
導語
發(fā)育機制的再激活發(fā)生在疾病發(fā)病機制和組織修復期間。因此,了解正常發(fā)育下細胞的可塑性以及局部細胞信號環(huán)境在決定細胞命運和功能中的作用,可能為先天性肺病、慢性疾病發(fā)病機制和細胞對治療的反應提供關(guān)鍵見解。在這里,我們構(gòu)建了一個發(fā)育中的人類胎兒肺的綜合轉(zhuǎn)錄組圖譜,該圖譜捕獲了超過 150000個胎兒肺細胞,并確定了揭示上皮區(qū)室內(nèi)顯著細胞可塑性的發(fā)育軌跡。通過空間分辨轉(zhuǎn)錄組學,我們確定了可能決定發(fā)育中肺內(nèi)細胞命運和行為的推定時間調(diào)節(jié)的細胞信號傳導相互作用。最后,我們揭示了hPSC的分化沿著相似的發(fā)育軌跡進行,并產(chǎn)生與在人胎兒肺上皮中觀察到的相似的胎兒細胞類型/狀態(tài)。
影響細胞命運決定的發(fā)育過程是由細胞微環(huán)境中精確的細胞間信號傳導相互作用介導的。雖然細胞可能共享轉(zhuǎn)錄組基因特征,但細胞自主或非細胞自主信號轉(zhuǎn)導可能決定這些細胞如何發(fā)育。在這里,我們專注于上皮-上皮細胞和上皮-基質(zhì)細胞之間的信號傳導通訊。然而,它并不排除免疫、內(nèi)皮和神經(jīng)(雪旺)細胞的貢獻,這些細胞也是共同發(fā)育并居住在前兩種細胞類型附近。事實上,免疫細胞群對上皮發(fā)育的貢獻最近已在胎肺中得到證實。相應地,我們在上皮細胞的軌跡分析中看到了白細胞介素通路的表達,這可能表明來自免疫細胞的信號傳導。在我們的細胞通訊和軌跡分析中,我們顯示了靶向 TP 細胞的FGF、WNT和 NOTCH信號通路的時間變化。信號轉(zhuǎn)導的空間和時間調(diào)節(jié)都可以改變細胞錯綜復雜的微環(huán)境,并影響其行為和潛在的細胞命運變化。為了確認我們的結(jié)果,使用Xenium來優(yōu)化上皮細胞和基質(zhì)細胞類型的空間接近度,并通過TP分析了推定的信號轉(zhuǎn)導。
我們研究的一個重要方面是 hPSC 衍生的胎肺分化的基準。在這里,我們表明 hPSC 衍生的胎兒肺細胞和類器官捕獲了在天然組織中觀察到的幾種胎兒特異性細胞類型/狀態(tài)和分化軌跡。具體來說,我們表明分化的 hPSC 衍生的胎兒肺細胞主要代表增殖細胞和祖細胞,而 hPSC 衍生的胎兒類器官包含代表肺發(fā)育后期的分化細胞。由于并非所有細胞類型和軌跡都在 hPSC 分化中被捕獲,因此未來的研究可能會利用 LR 相互作用的預測和特定信號通路的富集來修改當前的分化方案并改善體外其他上皮細胞類型的產(chǎn)生。總體而言,我們的工作支持使用 hPSC 衍生的胎肺上皮模型作為研究胎肺上皮譜系發(fā)育的潛在替代模型。重要的是,hPSC 分化是實驗上易于處理的模型,可用于研究更廣泛的發(fā)育范圍和疾病的胎兒起源,這使得這些模型的基準測試和驗證尤為重要。
總之,我們的研究確定了有助于早期人類胎兒肺上皮動態(tài)發(fā)育的細胞類型、狀態(tài)、軌跡以及局部細胞和信號相互作用。了解人類胎兒肺細胞多樣性及其在發(fā)育中的作用將改進當前 hPSC 的分化方案,從而為未來先天性和慢性肺病的治療生成更好的類器官模型或真正的細胞類型。
參考文獻:
Quach H, Farrell S, Wu MJM, Kanagarajah K, Leung JW, Xu X, Kallurkar P, Turinsky AL, Bear CE, Ratjen F, Kalish B, Goyal S, Moraes TJ, Wong AP. Early human fetal lung atlas reveals the temporal dynamics of epithelial cell plasticity. Nat Commun. 2024 Jul 13;15(1):5898. doi: 10.1038/s41467-024-50281-5. PMID: 39003323; PMCID: PMC11246468.